


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE EFFECT OF STEM ROT AT COMPOSITION AND CONTENT OF PHENOLIC COMPOUNDS IN LEAVES OF BIRCH (BETULA PENDULA ROTH.) AT THE FORESTS OF THE URALS UNDER ANTHROPOGENIC INFLUENCE

Как известно, одной из важных проблем в лесном хозяйстве Урала являются инфекционные болезни древесного яруса. Вследствие многократного порослевого возобновления и интенсивного антропогенного воздействия пораженность гнилевыми болезнями березняков в лесостепи Зауралья очень высока (60-85%) [4,5].
В целом, несмотря на интенсивные исследования в этом направлении, биохимические аспекты резистентности к гнилевым болезням и взаимодействия в системе: «патоген-растение-хозяин» остаются недостаточно выясненными, так как одни авторы отмечают накопление фенольных соединений в зоне некроза, вызванного инокуляцией патогенов, другие – их снижение [6-8,12].
Наиболее распространенным подходом в исследованиях является изучение биохимических процессов при искусственной инокуляции патогенов в зоне инокуляции. Ряд авторов отмечает, что некоторые фенольные соединения обладают выраженной фунгицидной активностью [10,11]. Имеются также единичные сообщения о том, что микозы у древесных растений сопровождаются активизацией синтеза фенольных соединений, детерминирующих параметры конститутивной резистентности [9]. Таким образом, в целом имеющиеся данные противоречивы. Исходя из этого, нами проведены исследования влияния поражения березы повислой стволовыми гнилями на состав и содержание фенольных соединений в листьях.
Материалы и методы
Для хроматографического анализа проводился сбор листьев березы повислой в Каменск-Уральском р-не Свердловской обл. Пробные площади располагались в березняках вблизи пос. «Покровское». Взятие каждой пробы осуществлялось от нескольких деревьев. Затем пробы смешивались для получения средней пробы. Непосредственно перед отбором проб определяли наличие или отсутствие пораженности дерева стволовыми и корневыми гнилями и стадию развития болезни с помощью взятия кернов из ствола и 3 корневых лап и их анализа. Как показали исследования, в припоселковых березовых лесах средний уровень антропогенной трансформации высок (4 стадия) [2]. Основной источник антропогенного воздействия: рекреационная и пастбищная дигрессия. Контрольные пробы отбирали от здоровых деревьев, не пораженных гнилевыми и другими болезнями, опытные пробы брали с деревьев, идентифицированных нами как пораженные стволовыми гнилями. Отбор проб проводился на одних и тех же пробных площадях, от деревьев одинакового диаметра, растущих в одинаковых лесорастительных условиях. Сразу после сбора листья березы высушивали при 600С, затем размалывали. После этого навеску с 2 г размолотых листьев смешивали с 20 мл 95% этанола. Экстракцию фенольных соединений из листьев березы проводили в обратном холодильнике на водяной бане в течение 30 мин. при кипении раствора. Затем суспензию центрифугировали при 10000 g в течение 10 мин. Супернатант фильтровали через фильтр с диаметром пор 0,2 мкм. Хроматографию проводили на жидкостном хроматографе Shimadzu LC-20 со спектрофотометрическим УФ детектором. Детектирование элюента осуществляли одновременно на двух полосах поглощения: 254 и 360 нм на колонке PerfectSil Target ODS-35мкм с обращенной фазой.
Градиентное элюирование проводилось в диапазоне 10–50 % со скоростью 1 мл в мин. при температуре 400С. Элюент А – ацетонитрил – 0,05 М фосфатный буферный раствор (рН=3,0); элюент В – ацетонитрил-вода (9:1). Продолжительность хроматографического анализа – 45 мин при 4000С. Из них от 0 до 30 мин. проводилось градиентное элюирование в диапазоне 10-50%, затем в течение 15 мин. при концентрации 50%. Для идентификации фенольных соединений использовали вещества-свидетели фирмы: Fluka, Sigma, Aldrich: аскорбиновая к-та, арбутин, галловая к-та, гиперозид, гидрохинон, кверцетин, изокверцетин, кофейная к-та, мирицетин, рутин, салицин, кафтаровая к-та, рутин; феруловая к-та, 3-КФК, 4-КФК, 5-КФК, кемпферол.
Результаты
Градиентный хроматографический анализ (ВЭЖХ) образцов из листьев березы повислой, взятых как с деревьев, пораженных стволовой гнилью, так и здоровых, позволил выявить 57 фенольных соединений (рис. 1).
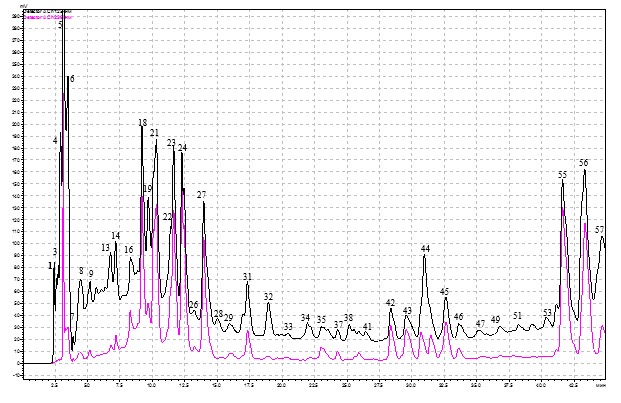
Рис. 1. ВЭЖХ фенольных соединений листьев березы повислой, пораженных стволовой гнилью. По оси абсцисс- tr (время удерживания), ординат-оптическая плотность
Примечания: 1 – аскорбиновая кислота; 4 – арбутин, 5 – галловая кислота; 7 – кафтаровая кислота; 8 – салицин, 13 – 4-кофеоилхинная кислота; 18 – рутин, 21 – 5-кофеоилхинная кислота; 23 – гиперозид; 24 – изокверцетин; 31 – мирицетин; 40 – кверцетин; 43 – кемпферол
Из общего количества фракций нами идентифицировано 13 фенольных соединений. Список идентифицированных фенольных соединений в листьях березы представлен в табл. 1. По химическому составу это были флавоноиды, фенолгликозиды, фенолкарбоновые и гидроксикоричные кислоты.
Для анализа основных, доминирующих тенденций влияния стволовой гнили на содержание фенольных соединений в листьях березы повислой нами проведен как тотальный сравнительный количественный попарный анализ основных выявленных при хроматографии пиков, так и сравнительный анализ идентифицированных нами фенольных соединений (табл. 1).
Таблица 1
Влияние стволовой гнили на состав и содержание фенольных соединений в листьях березы повислой
|
№ п/п |
Наименование |
Контроль Абсорбция (254 нм), mv |
Стволовая гниль Абсорбция (254 нм), mv |
Изменение содержания соединений (в % к контр.) |
Соотношение Y360 нм/ Y 254 нм |
ВУ |
|
1 |
Аскорбиновая к-та |
110,09 |
101,0 |
-8,18 |
0,0008 |
2,64 |
|
2 |
Арбутин |
285,0 |
193,0 |
-32,28 |
0,075 |
3,06 |
|
3 |
Галловая к-та |
90,5 |
237,0 |
+161,88 |
0,021 |
3,6 |
|
4 |
Салицин |
58,3 |
48,0 |
-17,24 |
0,0001 |
4,29 |
|
5 |
Кафтаровая к-та |
24,5 |
12,1 |
-50,61 |
0,095 |
4,003 |
|
6 |
4-кофеоилхинная к-та |
29,01 |
46,0 |
+36,93 |
0,30 |
6,7 |
|
7 |
Рутин |
43,0 |
113,0 |
+61,95 |
0,98 |
9,46 |
|
8 |
5-кофеоилхинная к-та |
90,06 |
143,0 |
+58,0 |
0,65 |
10,0 |
|
9 |
Гиперозид |
293,37 |
177,5 |
-39,50 |
0,884 |
10,93 |
|
10 |
Изокверцетин |
39,75 |
140,0 |
+252,2 |
1,00 |
11,74 |
|
11 |
Мирицетин |
22,5 |
44,2 |
+96,44 |
1,04 |
17,05 |
|
12 |
Кверцетин |
6,0 |
9,1 |
+51,66 |
0,97 |
25,4 |
|
13 |
Кемпферол |
21,8 |
20,5 |
--- |
1,22 |
28,5 |
Тотальный попарный сравнительный анализ образцов листьев пораженных и контрольных древостоев (пиков с идентичным ВУ и спектральным соотношением (S360 /S254)) убедительно продемонстрировал, что эти образцы значительно отличались между собой. Так, у 35,9% фракций их содержание заметно увеличивалось, у такого же количества уменьшалось, у 28,2% не изменялось. Для более глубокого анализа биохимических механизмов, происходящих в клетках листьев березы в насаждениях, пораженных стволовой гнилью, было целесообразно проанализировать тенденции реакций идентифицированных нами на хроматограммах фенольных соединений, так как функции ряда из них хорошо известны.
Как показали результаты исследований, у 50% идентифицированных нами фенольных соединений их содержание заметно увеличивалось. Это галловая к-та (161,88%), 4-кофеоилхинная к-та (36,93%), рутин (61,95%), 5-кофеоилхинная к-та (58,0%), изокверцетин (252,2%), мирицетин (96,44%) и кверцетин (51,66%). Обращает внимание значительный уровень возрастания активности галловой к-ты, изокверцетина и мирицетина. Одновременно снижается уровень активности аскорбиновой к-ты, арбутина, салицина, кафтаровой к-ты, гиперозида. Сравнение полученных нами результатов с имеющимися в литературе не дает положительных результатов, так как нами изучались биохимические изменения на значительной стадии пораженности, когда площадь пораженной древесины превышала 30% по диаметру, тогда как другие авторы изучали чаще биохимический состав после непосредственной инокуляции патогена. Так, синтез таких фенолгликозидов, как арбутин и салицин не активируется, но, в целом общий уровень активности идентифицированных соединений значительно возрастает.
Учитывая то, что большинство идентифицированных нами фенольных соединений являются активными антиоксидантами, можно заключить, что биохимические процессы борьбы с оксидативным стрессом у березы, пораженной стволовой гнилью, заметно активизируются. Вместе с тем, известно, что некоторые патогенные грибы сами выделяют антиоксиданты для разрушения Н2О2 и супероксидного радикала, которые выполняют защитную функцию, способствующую выживанию патогена (Абрамова и др., 2008). Поэтому возможно, что столь заметное увеличение активности ряда фенольных соединений, обладающих высокой антиоксидантной активностью, обусловлено защитной реакцией дерева-хозяина на заражение гнилевыми болезнями. Вместе с тем, мы предполагаем, что это одна из дополнительных функций фенольных соединений. Кроме того, обращает внимание ингибирование синтеза фенолгликозидов (арбутина, салицина) в насаждениях, пораженных стволовыми гнилями, детерминирующих параметры индуцированной энтоморезистентности. Полученные результаты предполагают, что развитие гнилевых болезней у березы повислой может сопровождаться снижением уровня энтоморезистентности. Это хорошо соответствует полученным нами ранее результатам о том, что поражение стволовыми гнилями березы сопровождается заметным снижением энтоморезистентности и возрастанием степени дефолиации крон непарным шелкопрядом, по сравнению со здоровыми деревьями (Колтунов, Клобуков, 2009).
Рецензенты:
Усольцев В.А., д.с.-х.н., профессор, профессор Уральского государственного лесотехнического университета, г. Екатеринбург;
Менщиков С.Л., д.с.-х.н., заведующий лабораторией экологии техногенных растительных сообществ ФГБУН Ботанический сад Российской Академии наук, г. Екатеринбург.
Библиографическая ссылка
Колтунов Е.В., Яковлева М.И. ВЛИЯНИЕ СТВОЛОВОЙ ГНИЛИ НА СОСТАВ И СОДЕРЖАНИЕ ФЕНОЛЬНЫХ СОЕДИНЕНИЙ В ЛИСТЬЯХ БЕРЕЗЫ ПОВИСЛОЙ (BETULA PENDULA ROTH.) В ЛЕСАХ ЗАУРАЛЬЯ В УСЛОВИЯХ АНТРОПОГЕННОГО ВОЗДЕЙСТВИЯ // Современные проблемы науки и образования. 2015. № 5. ;URL: https://science-education.ru/en/article/view?id=21797 (дата обращения: 26.06.2026).