


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
EFFECT OF VARIOUS CONCENTRATIONS OF NITROGEN OXIDE DONOR SNP ON ZEIDEL ASCITES HEPATOMA CELLS SURVIVAL IN VITRO

В настоящее время одну из важных ролей в регрессии опухолей отводят активным формам кислорода и азота. Оксид азота и его метаболиты являются сильным цитотоксическим оружием иммунной системы, оказывающим как прямое повреждающее действие на клетки, так и косвенное, путем регуляции клеточного цикла и процессов клеточной гибели.
Оксид азота (NO) является важным регуляторным фактором, участвующим в индукции апоптоза [4], пролиферации клеток [2] и тонуса сосудов [3]. В то же время активные метаболиты азота наряду с активными формами кислорода способны повреждать различные клеточные структуры, изменяя их функциональную активность. Так, процесс нитрозилирования белков под действием активных форм азота является одним из механизмов регуляции активности некоторых ферментных систем [7]. Следует отметить, что некоторые метаболиты NO, такие как S-нитрозотиолы и сам NO, при определённых условиях могут проявлять антиоксидантную активность [5]. Например, оксид азота может связываться с ионами железа, которые катализируют реакции свободно-радикального окисления.
Различные исследования показали роль NO в индукции генотоксического повреждения, а также его участие в продвижении опухоли и прогрессии от посреднических важных процессов, в том числе ангиогенеза, роста опухолевых клеток, и инвазии [6]. При этом оксид азота является важным компонентом иммунной реакции при росте некоторых типов опухолей, например, защищает от рака толстой кишки у мышей [10]. Высокие уровни NO обладают сильной генотоксичностью через образование канцерогенных нитрозаминов или путём повреждения систем репарации ДНК [8]. Однако NO также способен и уменьшать воздействие активных форм кислорода на ДНК [9].
Асцитная гепатома Зайдела является перевиваемой культурой и при её трансплантации в брюшную полость развивается асцитная форма опухоли, которая представляет собой клеточные комплексы и отдельные клетки, растущие в сильно геморрагической асцитической жидкости. Эта культура является удобной моделью исследования действия противоопухолевых препаратов на рост опухоли в организме. Однако доля исследований, посвященных изучению изолированных клеток данной опухоли in vitro в настоящее время незначительна. В данной работе было исследовано действие различных концентраций экзогенного донора оксида азота SNP на выживаемость клеток асцитной гепатомы Зайдела, культивируемых в питательной среде.
Материалы и методы исследования.
Для формирования опухоли использовали клетки асцитной гепатомы Зайдела, хранящиеся в криобанке. Клетки трансплантировали в брюшную полость крысы путем введения 0,8 мл клеток в количестве 1•107 кл/мл. Образовавшеюся асцитную жидкость отбирали на 5 сутки после трансплантации, центрифугировали при 400g в течение 10 мин., затем отмывали 2 раза в растворе NaCl (0,9%). Отмытые клетки наслаивали на градиент Percoll – NaCl c плотностями 1,04, 1056 и 1,078 г./мл. и центрифугировали 40 мин при 4000g. Далее отбирали шприцом слой опухолевых клеток на границе плотности 1,056 г./мл., Фракцию клеток промывали 2 раза средой DMEM. Эксперименты проводили в культуральных планшетах в среде DMEM c 10% эмбриональной телячьей сывороткой при 37°C, концентрация CO2 - 5%. Объем среды культивирования 0,7 мл, начальная концентрация клеток 106 кл./мл. Подсчет клеток проводили в камере Горяева. Определение жизнеспособности клеток осуществляли по окраске трипановым синим. Клеточный цикл регистрировали с помощью проточной цитометрии с окрашиванием пропидиумиодитом (PI). Для этого клетки гепатомы Зайдела фиксировались 70% С2Н5OH. Пробы центрифугировали 10 минут при 400g, удаляли спиртовую фракцию и добавляли 500мкл 0,0035% раствора (Pl) и 500 мкл 0,0035% раствора РНКазы в фосфатном буфере. Пробы инкубировали в термостате 30 минут при температуре 37°C. Затем проводили исследование на проточном цитофлюориметре (Partec PAS III, Германия)
Результаты исследования и их обсуждение
Для проведения исследования опухолевые клетки отбирались из асцитной жидкости экспериментального животного на II фазе развития экспериментальной опухоли. В этот период наблюдается интенсивный рост популяции клеток гепатомы Зайдела, а патологический процесс в организме не перешёл в необратимую фазу и, следовательно, ещё актуально проведение противоопухолевой терапии [1]. Выделенные клетки являются переживающей культурой и длительное их культивирование in vitro невозможно. В связи с этим эксперимент ограничен 24ч.
На рис. 1 представлены кривые роста асцитной гепатомы Зайделя в присутствие различных концентраций донора оксида азота SNP. Как видно из рисунка в процессе культивирования контрольных клеток гепатомы (в отсутствие SNP) в течение первых 12ч. наблюдается рост концентрации опухолевых, сменяемый устойчивой гибелью клеток при дальнейшем культивировании. При внесении в среду культивирования источника оксида азота (нитропрусида натрия - SNP) был отмечен нелинейный эффект действия различных концентраций данного соединения. Так при концентрации SNP ~0,02 мМ через 12 часов культивирования наблюдается достоверное увеличение количества клеток гепатомы по сравнению с контрольным образцом. По-видимому, данный эффект обусловлен активирующим воздействием данной концентрации SNP на пролиферативную активность клеток. При дальнейшем культивировании клеток эффект SNP несколько снижается и к 24 часам концентрация клеток практически не отличалась от контрольных значений. При дальнейшем ступенчатом увеличении концентрации донора NO до 0,2мМ, 1мМ и 2мМ количество опухолевых клеток в единице объема дозозависимо снижалось, что свидетельствует о цитотоксическом эффекте (рис 1, кривые 3,4 и 5).
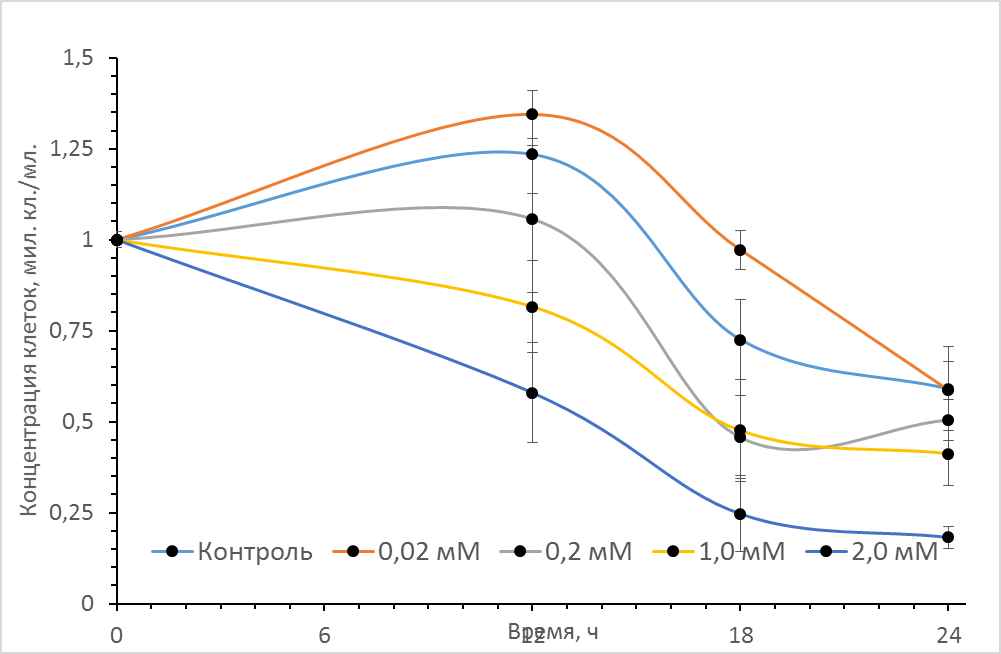
Рис. 1 Влияние различных концентраций донора оксида азота (SNP) на рост клеток асцитной гепатомы Зайдела. Культивирование проводилось в культуральных планшетах в среде DMEM c 10% эмбриональной телячьей сывороткой при 37°C, концентрация CO2 - 5%. Объем среды культивирования 0,7 мл, начальная концентрация клеток 106 кл./мл. Аналогичные результаты были получены в пяти независимых экспериментах.
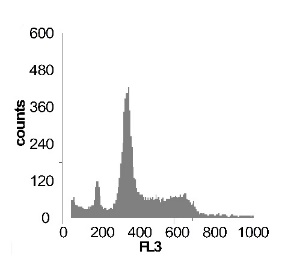
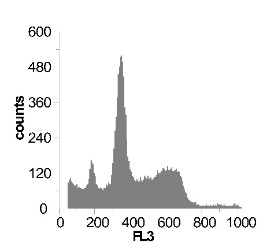
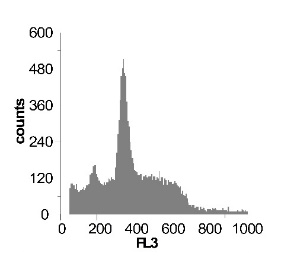
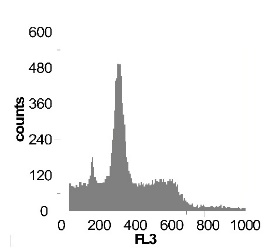
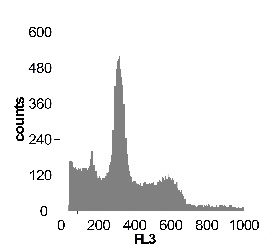
Используя метод проточной цитофлуориметрии, было исследовано состояние клеточного цикла опухолевых клеток по истечению 12 ч культивирования. Обнаружено, что с ростом концентрации SNP в среде культивирования увеличивается sub-G1 область на гистограмме клеточного цикла (рис 2), что говорит об увеличение фрагментации ДНК, и как следствие, гибели клеток. Это свидетельствует о цитотоксической активности SNP по отношению к клеткам гепатомы, обусловленной способностью оксида азота индуцировать апоптоз.
Наряду с цитотоксическим воздействием SNP на клетки гепатомы обнаружено, что при концентрации SNP ~ 0,02 мМ увеличивается доля клеток, находящихся в S и G2/M фазах клеточного цикла, что говорит об активации клеточного деления. Это увеличение пролиферативной активности не только компенсирует цитотоксический эффект SNP, но и позволяет увеличить популяцию клеток гепатомы в течении первых 12-18 ч культивирования. Дальнейшее увеличение концентрации донора оксида азота до 0,2мМ приводит к увеличению доли клеток в предмитотической фазе S и снижению в G2/M. В результате этого цитотоксический эффект уже не полностью компенсируется, что подтверждается снижением выживаемости клеток гепатомы Зайдела при культивировании (рис. 2) Дальнейшее увеличение концентрации SNP приводит к увеличению гибели клеток, причем, в основном, за счет находящихся в S и G2/M фазах клеточного цикла.
|
Контроль |
SNP |
|||
|
|
|
|
|
|
|
- |
0,02мМ |
0,2мМ |
1,0мМ |
2,0мМ |
|
|
|
|
|
|
|
|
контроль |
SNP 0,02 мМ |
SNP 0,20 мМ |
SNP 1,00 мМ |
SNP 2,00 мМ |
|
Sub-G1 |
16,7% |
19,7% |
22,6% |
23,3% |
28,9% |
|
G1 |
51,7% |
44,7% |
45,3% |
48,0% |
45,6% |
|
S |
13,0% |
13,6% |
16,0% |
13,0% |
11,1% |
|
G2/M |
18,6% |
22,0% |
16,1% |
15,7% |
14,4% |
Рис. 2 Анализ клеточного цикла клеток асцитной гепатомы Зайдела, инкубированных в присутствии SNP с концентрациями 0,02 мМ ; 0,2 мМ ; 1,0 мМ ; 2,0 мМ. Клетки окрашивали PI после 12 часов культивирования. Клеточный цикл представлен как процент клеточной популяции в различных фазах (sub-G 1; G1; S; G2/M) За мертвые клетки принимали sub-G1 область гистограммы. Аналогичные результаты были получены в пяти независимых экспериментах.
На основании полученных данных можно сделать вывод, что концентрация донора оксида азота SNP больше 0,2мM оказывает цитотоксический эффект по отношению к клеткам гепатомы Зайдела в условиях in vitro. При этом в значительной мере снижется доля делящихся клеток. Подобный эффект является поводом для использования доноров оксида азота как противоопухолевого агента в комплексной терапии онкологических заболеваний. Однако надо учитывать, что концентрации SNP ниже 0,2 мМ, напротив, увеличивают пролиферации клеток и, следовательно, способствуют расту опухоли.
Рецензенты:
Новоселова Е.Г., д.б.н., профессор, ведущий научный сотрудник лаборатории механизмов рецепции ФГБУН Института биофизики клетки РАН, г. Пущино;
Асланиди К.Б., д.ф.-м.н., ведущий научный сотрудник лаборатории биофизики внутриклеточной регуляции ФГБУН Института теоретической и экспериментальной биофизики РАН, г. Пущино.
Библиографическая ссылка
Куприянова Е.С., Наумов А.А., Поцелуева М.М., Серебрякова Л.Т. ВЛИЯНИЯ РАЗЛИЧНЫХ КОНЦЕНТРАЦИЙ ДОНОРА ОКСИДА АЗОТА SNP НА ВЫЖИВАЕМОСТЬ КЛЕТОК АСЦИТНОЙ ГЕПАТОМЫ ЗАЙДЕЛА IN VITRO // Современные проблемы науки и образования. 2014. № 6. ;URL: https://science-education.ru/en/article/view?id=16119 (дата обращения: 02.08.2026).