


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE MODEL OF GROWTH ONE-TYPE POPULATION IN SPACE AND TIME

Введение
При исследовании популяций, развивающихся в пространстве и времени, возникает задача оценки численности популяции в произвольных областях в некоторый момент времени, зная начальное распределение [1]. Под однотипной популяцией в данной работе рассматривается популяция бактерий. Предполагается, что наблюдается не вся область, а лишь выборочные ее сектора в случайные моменты времени. При этом наблюдение сильно зашумлено – в момент подсчета числа бактерий в некоторой области бактерии могут перемещаться в соседние области. По этой причине разработка алгоритмов и математических моделей, предназначенных для анализа роста однотипных популяций, представляет актуальную научную проблему. Целью данной работы является построение адекватной математической модели, позволяющей анализировать развитие однотипной популяции в пространстве и времени. Методом исследования является имитационное стохастическое моделирование. Оценка реального числа бактерий в секторе на основе наблюдений осуществляется с помощью дискретной схемы Калмана.
Описание модели
Рассмотрим математическую модель роста однотипной популяции, развивающуюся из исходной популяции во времени и двумерном пространстве. Для определенности опишем процесс, являющийся моделью роста популяций бактерий.
Предположим, что ареал ![]() имеет форму прямоугольника. Разобьем ареал на одинаковые прямоугольные области
имеет форму прямоугольника. Разобьем ареал на одинаковые прямоугольные области![]() ,
, ![]() и
и ![]() ,
,![]() и
и![]() – натуральные числа, таким образом
– натуральные числа, таким образом ![]() ,
, ![]() и
и ![]() .
.
В каждой области ![]() ,
, ![]() и
и ![]() может находиться неотрицательное целое число бактерий. Каждая бактерия может с вероятностью
может находиться неотрицательное целое число бактерий. Каждая бактерия может с вероятностью ![]() (
(![]() ) поделиться на две. Обозначим
) поделиться на две. Обозначим ![]() процесс, считающий количество появившихся бактерий в результате деления внутри области
процесс, считающий количество появившихся бактерий в результате деления внутри области ![]() (
(![]() и
и ![]() ). Предполагается, что интенсивность деления пропорциональна количеству бактерий в области в момент времени
). Предполагается, что интенсивность деления пропорциональна количеству бактерий в области в момент времени ![]() с коэффициентом пропорциональности
с коэффициентом пропорциональности ![]() , т.е. компенсатор процесса
, т.е. компенсатор процесса ![]() описывается классической моделью Мальтуса [3] и равен:
описывается классической моделью Мальтуса [3] и равен:
![]() (1)
(1)
где ![]() – количество бактерий в момент времени t внутри области
– количество бактерий в момент времени t внутри области ![]() (
(![]() и
и ![]() ). Основные определения и термины для описания семи мартингалов см. в [2].
). Основные определения и термины для описания семи мартингалов см. в [2].
Рассмотрим также процессы перехода бактерий в область ![]() из соседних областей и ухода бактерии из области
из соседних областей и ухода бактерии из области ![]() в соседние области. Обозначим их
в соседние области. Обозначим их ![]() и
и ![]() (
(![]() и
и ![]() ) соответственно. Предполагается, что компенсатор процесса ухода
) соответственно. Предполагается, что компенсатор процесса ухода ![]() бактерии из области
бактерии из области ![]() (
(![]() и
и ![]() ) также пропорционален количеству бактерий в области в момент времени
) также пропорционален количеству бактерий в области в момент времени ![]() с коэффициентом пропорциональности
с коэффициентом пропорциональности ![]() .
.
где ![]() – интенсивность ухода бактерии из области из области
– интенсивность ухода бактерии из области из области ![]() (
(![]() и
и ![]() ).
).
а) 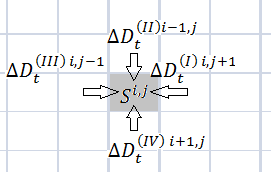 б)
б) 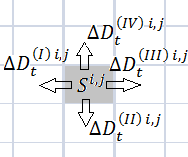
Рисунок 1. Переходы бактерий из области ![]() и в область
и в область ![]() из соседних областей
из соседних областей
Обозначим ![]() – количество бактерий, перешедших из области
– количество бактерий, перешедших из области ![]() в область
в область ![]() ,
, ![]() и
и ![]() в момент времени
в момент времени ![]() ;
; ![]() – из области
– из области ![]() в область
в область ![]() ,
, ![]() и
и ![]() ;
; ![]() – из области
– из области ![]() в область
в область ![]() ,
, ![]() и
и ![]() и
и ![]() – из области
– из области ![]() в область
в область ![]() ,
, ![]() и
и ![]() , см. рисунок 1 (а, б). Пусть бактерии не умирают, тогда выполняется соотношение:
, см. рисунок 1 (а, б). Пусть бактерии не умирают, тогда выполняется соотношение:
![]() (3)
(3)
Предполагается, что бактерия не может одновременно перейти в две разных области, т. е. выполняется равенство (4).
![]() P-п. н. (4)
P-п. н. (4)
Предположим, что переходы бактерий из области ![]() будут происходить в момент скачков пуассоновского процесса
будут происходить в момент скачков пуассоновского процесса![]() . Обозначим эти моменты
. Обозначим эти моменты ![]() ,
, ![]() ,
,![]() и
и ![]() . Тогда вероятности переходов будут равны:
. Тогда вероятности переходов будут равны:
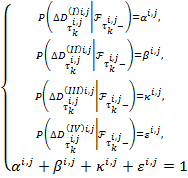 (5)
(5)
С учетом (5) процесс ![]() равен
равен
![]() (6)
(6)
Предположим, что ареал замкнут всюду, т. е. ![]() ;
; ![]() ;
; ![]() и
и ![]() .
.
С учетом формул (1-3, 6) получаем, что количество бактерий в области ![]() (
(![]() и
и ![]() ) в момент времени t равно
) в момент времени t равно
![]() (7)
(7)
где ![]() – начальное количество бактерий в из области
– начальное количество бактерий в из области ![]() (
(![]() и
и ![]() ).
).
Процесс наблюдения ![]() за процессом
за процессом ![]() будет выглядеть следующим образом
будет выглядеть следующим образом
![]() ,
, ![]() (8)
(8)
![]() ,
, ![]() ,
,
где ![]() – целочисленная случайная величина, распределенная по закону Пуассона с параметром
– целочисленная случайная величина, распределенная по закону Пуассона с параметром ![]() , ошибка измерений.
, ошибка измерений.
Предположим, что распределение бактерий в начальный момент времени по областям ареала ![]() неравномерно. Требуется по наблюдению
неравномерно. Требуется по наблюдению ![]() оценить численность бактерий в каждой области ареала.
оценить численность бактерий в каждой области ареала.
Теоретическая оценка математического ожидания числа бактерий
Математическое ожидание процесса ![]() , считающего количество бактерий в области
, считающего количество бактерий в области ![]() (
(![]() и
и ![]() ), из уравнений (1, 2, 6, 7) равно:
), из уравнений (1, 2, 6, 7) равно:
![]()
![]()
![]()
![]() , (9)
, (9)
где ![]() и
и ![]() , причем слагаемые с индексами: 0,
, причем слагаемые с индексами: 0, ![]() и
и ![]() равны 0. Запишем полученные уравнения (9) в дифференциальном виде:
равны 0. Запишем полученные уравнения (9) в дифференциальном виде:
![]()
![]() (10)
(10)
Получена система ![]() однородных линейных дифференциальных уравнений (10) с
однородных линейных дифференциальных уравнений (10) с ![]() неизвестными и начальными значениями
неизвестными и начальными значениями ![]() . Решение таких систем подробно изложено в [4]. Решение в общем виде можно записать через матричную экспоненту:
. Решение таких систем подробно изложено в [4]. Решение в общем виде можно записать через матричную экспоненту:
![]() (11)
(11)
где ![]() – вектор,
– вектор, ![]() ;
; ![]() – матрица, составленная из коэффициентов уравнений (10);
– матрица, составленная из коэффициентов уравнений (10); ![]() – начальное значение вектора
– начальное значение вектора![]() .
.
Оценка численности числа бактерий
Предположим, что наблюдение за областями ![]() и
и ![]() происходит в случайные моменты времени
происходит в случайные моменты времени ![]() ,
, ![]() скачков пуассоновского процесса
скачков пуассоновского процесса ![]() с интенсивностью
с интенсивностью ![]() одновременно во всех областях
одновременно во всех областях ![]() . И предположим также, что при подсчете количества бактерий в области
. И предположим также, что при подсчете количества бактерий в области ![]() и
и ![]() происходит ошибка
происходит ошибка ![]() , т. е.
, т. е.
![]() (11)
(11)
где ![]() ,
, ![]() – совокупность независимых случайных величин, c нулевым средним и дисперсией
– совокупность независимых случайных величин, c нулевым средним и дисперсией ![]() .
.
Значение процесса![]() в моменты времени
в моменты времени ![]() будет отличаться от математического ожидания
будет отличаться от математического ожидания ![]() на некоторую мартингальную часть. В предыдущем пункте был рассмотрен способ, позволяющий отыскать теоретическую оценку математического ожидания числа бактерий. Само решение зависит от конкретных значений интенсивностей переходов
на некоторую мартингальную часть. В предыдущем пункте был рассмотрен способ, позволяющий отыскать теоретическую оценку математического ожидания числа бактерий. Само решение зависит от конкретных значений интенсивностей переходов ![]() и
и ![]() и весовых коэффициентов
и весовых коэффициентов![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() , поэтому обозначим, что от момента
, поэтому обозначим, что от момента ![]() до момента
до момента ![]() математическое ожидание числа бактерий меняется по закону:
математическое ожидание числа бактерий меняется по закону:
![]() (12)
(12)
где ![]() – некоторая функция, которая ищется исходя из решений уравнений (10). Суммируя вышесказанное, получаем формулу (13):
– некоторая функция, которая ищется исходя из решений уравнений (10). Суммируя вышесказанное, получаем формулу (13):
![]() , (13)
, (13)
где ![]() – независимые одинаково распределенные случайные величины с нулевым средним и дисперсией
– независимые одинаково распределенные случайные величины с нулевым средним и дисперсией ![]() .
.
Рассмотрим способ оценки реального числа бактерий ![]() по наблюдению за областью
по наблюдению за областью ![]() и
и ![]() на основе дискретной схемы Калмана [5].
на основе дискретной схемы Калмана [5].
В каждый момент времени оценка реального числа бактерий в области ![]() будет считаться как средневзвешенное между теоретической оценкой бактерий и наблюдаемыми значениями с коэффициентами
будет считаться как средневзвешенное между теоретической оценкой бактерий и наблюдаемыми значениями с коэффициентами ![]() и
и ![]() соответственно, где
соответственно, где ![]() – коэффициенты Калмана [5].
– коэффициенты Калмана [5].
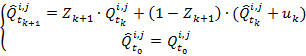 (14)
(14)
где ![]() – коэффициент Калмана,
– коэффициент Калмана, ![]() . Нужно так подобрать коэффициенты
. Нужно так подобрать коэффициенты![]() , чтобы получившаяся оценка числа бактерий
, чтобы получившаяся оценка числа бактерий ![]() была наиболее близкой к реальному числу
была наиболее близкой к реальному числу ![]() . К примеру, если известно, что ошибка обнаружения при подсчете числа бактерий очень мала, то коэффициенту
. К примеру, если известно, что ошибка обнаружения при подсчете числа бактерий очень мала, то коэффициенту ![]() нужно придать больший вес. И наоборот, если известно, что значение числа бактерий при подсчете очень не точное, следует больше ориентироваться на теоретически предсказанное значение
нужно придать больший вес. И наоборот, если известно, что значение числа бактерий при подсчете очень не точное, следует больше ориентироваться на теоретически предсказанное значение ![]() .
.
Если обозначить ошибку ![]() между реальным значением числа бактерий
между реальным значением числа бактерий ![]() в области
в области ![]() и их оценкой
и их оценкой ![]() , тогда задачу оценивания можно переформулировать в задачу минимизации среднеквадратической ошибки в момент времени
, тогда задачу оценивания можно переформулировать в задачу минимизации среднеквадратической ошибки в момент времени ![]() , см. формулу (16).
, см. формулу (16).
![]() (15)
(15)
![]() (16)
(16)
Теорема. При сделанных выше предположениях оценка числа бактерий ![]() будет оптимальной при:
будет оптимальной при:
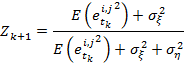
Доказательство
Используя уравнения (11), (13) и (14), перепишем уравнение (15) в виде:
![]()
![]()
![]()
![]()
Из предположения независимости случайных величин, входящих в ![]() и с учетом того, что
и с учетом того, что ![]() и
и ![]() получаем, что выражение (15) можно переписать в виде:
получаем, что выражение (15) можно переписать в виде:
![]()
Это выражение принимает минимальное значение при
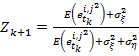 (17)
(17)
Среднеквадратичная ошибка, минимизирующая значения коэффициентов Калмана, при этом равна:
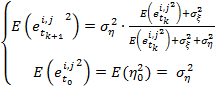 (18)
(18)
Что и требовалось доказать.
Заключение
Рассмотренная в статье модель позволяет описать рост однотипной популяции в пространстве и времени на основе семимартингальных точечных процессов по известному начальному распределению. Получена итерационная формула (17) для вычисления оптимальной оценки числа бактерий в произвольной области на основе нескольких наблюдений.
Работа выполнена при поддержке Министерства образования и науки РФ (НИР, проводимые в рамках государственного задания Министерства образования и науки РФ на 2013 г., Программы стратегического развития УлГУ на 2012–2016 гг., Программы развития деятельности студенческих объединений УлГУ на 2012-2013 гг.).
Рецензенты:
Кемер А.Р., д.ф.-м.н., профессор, ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск.
Андреев А.С., д.ф.-м.н., профессор, ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск.
Библиографическая ссылка
Бутов А.А., Егоров А.Г. МОДЕЛЬ РОСТА ОДНОТИПНОЙ ПОПУЛЯЦИИ В ПРОСТРАНСТВЕ И ВРЕМЕНИ // Современные проблемы науки и образования. 2013. № 6. ;URL: https://science-education.ru/en/article/view?id=10794 (дата обращения: 01.07.2026).