



Проблема исследования механизмов и факторов популяционной динамики лесных насекомых-филлофагов, образующих периодические крупномасштабные и пандемические вспышки массового размножения, является одной из наиболее важных в популяционной экологии и общей биологии. Это обусловлено необходимостью как разработки более эффективных математических моделей популяционной динамики, так и поиска методов управления популяциями, так как многие из этой группы насекомых-фитофагов способны наносить значительный эколого-экономический ущерб лесным фитоценозам. К настоящему времени основные периоды вспышек массового размножения лесных насекомых-филлофагов изучены достаточно детально [4; 15; 16; 22]. Но остаются недостаточно изученными как механизмы переключения циклов, так и группа скрытых цикличностей, так как часть визуализированных периодов является следствием наложения исходных циклов. Для изучения этих проблем мы использовали спектральный анализ Фурье, позволяющий точно вычислить периодичность и мощность циклов, а также обнаружить скрытые цикличности. Учитывая хорошо известную взаимосвязь вспышек массового размножения с периодичностью наступления весенне-летних засух [1; 4; 15-17], мы также изучили сравнительные спектральные характеристики циклов засух и популяционной динамики различных географических популяций непарного шелкопряда на Урале.
К настоящему времени установлено, что временную организацию всех биологических систем характеризует целый набор различных периодов – спектр [5; 7; 14; 37]. На «надорганизменном» уровне внешним проявлением их являются биологические ритмы, например периодические изменения численности в популяции. Следовательно, динамика биологических систем характеризуется спектром периодов. Этот изоморфизм спектров может быть интерпретирован как согласование биологических ритмов с временной структурой внешней среды. Оно осуществляется «затягиванием» биологических автоколебаний «внешними датчиками времени», природа которых может быть самой разнообразной: фотопериодизм, температурный ритм и цикличность влажности, смена времен года, климатические циклы. Могут выступать синхронизующим сигналом и электромагнитные колебания [25].
Адаптированность популяции можно оценить и по ее хроноэкологической структуре. К любым внешним ритмам она может адаптироваться путем синхронизации («затягивание» собственного ритма). Таким образом, устойчивый популяционный цикл должен существовать сам, его механизмом являются внутрипопуляционные процессы, внешний же геофизический ритм выполняет функцию лишь синхронизирующего ритма [11]. В ином случае адаптация становится невозможной, так как при отсутствии собственного ритма заданной частоты отсутствует и возможность синхронизации к внешнему периодическому процессу. По мнению В.Б. Чернышева, биоритмы – это суть самого биологического процесса, одна из необходимых частей его механизма [30]. Оценка адаптационных возможностей популяции может быть сделана по числу устойчивых циклов, из суперпозиции которых складывается кривая изменений ее численности. Если популяция имеет множество ритмов численности, то у нее увеличиваются возможности адаптации к подходящим для нее циклическим изменениям внешней среды. Такие циклы в динамических процессах нетрудно выделить и определить их характеристики, что и было давно сделано для многих популяций животных [11; 12; 23; 24].
Для детального сравнительного спектрального анализа популяционной динамики различных географических популяций непарного шелкопряда необходимо преобразование имеющихся данных по временным рядам с временной шкалы на частотную. Тогда изменения численности будут выглядеть как спектр некоторого количества периодов (рис. 2).
Основной целью исследований был поиск и идентификация всех периодических составляющих многолетней динамики вспышек массового размножения непарного шелкопряда на Урале. Основными задачами являлись: определение спектра ритмов численности в разных частях ареала; расчет периодов и мощностей гармонических составляющих его динамики численности; поиск возможных внешних синхронизаторов для различных циклов в многолетней динамике вспышек массового размножения непарного шелкопряда.
Материал и методика
Объектом исследования были временные ряды многолетней динамики вспышек массового размножения различных географических популяций непарного шелкопряда. Это популяция в Челябинской области (за 62 года) [15; 16], Свердловской обл. (за 52 года) [15; 16], Башкортостане (за 208 лет) [22] и Оренбургской обл. (за 23 года) [28]. В качестве объекта для сравнения использовались данные по динамике вспышек массового размножения «европейской» географической популяции непарного шелкопряда (почти за 150 лет) [3]. Кроме того, анализировали многолетнюю динамику гидротермических коэффициентов Селянинова (апрель-сентябрь) в Свердловской и Челябинской обл.
Для выявления скрытых колебаний численности непарного шелкопряда был использован анализ временных рядов [8]. Эмпирически определяемыми параметрами были: шаг суммирования, длина автокорреляционной функции, форма и ширина корреляционного окна. Детальное описание методик расчета биологических ритмов дано А.А. Сорокиным [29] и Л.Н. Ердаковым [11]. В результате на спектре происходит визуализация распределения функции спектральной плотности, в каждой точке которой она соответствует средней мощности в полосе частот определенной ширины – «пик на спектре». Временные ряды популяционной динамики вспышек исследовались на наличие скрытых гармонических составляющих. Для каждого были построены спектры ритмов, а также рассчитаны периоды и мощности гармонических составляющих каждого такого спектра.
Результаты и их обсуждение
Спектральный анализ многолетней динамики вспышек массового размножения непарного шелкопряда в условиях Челябинской обл.
Как известно, за весь период наблюдений у этого фитофага было зарегистрировано пять
вспышек массового размножения, причем последняя, наиболее продолжительная, продолжается более семи лет (рис. 1). Интервал между первой и второй вспышками составлял 12 лет, между второй и третьей - 8 лет, между третьей и четвертой – 12 лет, а между четвертой и пятой – 24 года. Таким образом, очевидно, что, несмотря на относительно устойчивую цикличность засух (10-12 лет), периодичность вспышек массового размножения «челябинской» географической популяции непарного шелкопряда имеет как минимум 3 различных периода.
Как показали результаты спектрального анализа, на спектре идентифицируются 8-10 заметных гармоник (рис. 2). Одна из них, примерно 11-летняя (11,3), имеет максимальную мощность (табл. 1).Она является общеизвестной и идентифицируется многими авторами, как 10-12 летняя [4; 15; 16]. Очень близка к ней 8,2-летняя цикличность, меньшая по мощности. В низкочастотной области спектра выявлен второй по мощности, близкий к 40-летнему пик и более слабый – 18-летний. Первый из них близок к длительности некоторых внутривековых «брикнеровских» климатических циклов (35-45 лет) [32]. На их фоне развиваются циклы продолжительностью в 7-11 лет. Они описаны и другими авторами [10; 20]. Кроме этого периода, динамика вспышек массового размножения непарного шелкопряда имеет еще несколько различных по мощности скрытых периодичностей. Мы рассчитали значения периодов и мощностей этих гармонических составляющих (табл. 1). Третий по мощности – 3-4-летний цикл. Следующим является близкий к 5-летнему цикл. Он характерен для июль-августовских осадков [9] и даже может «затягиваться» соответствующим по величине георитмом Земли [33].
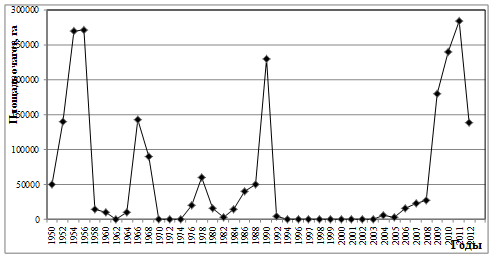
Рис. 1. Многолетняя динамика вспышек массового размножения непарного шелкопряда в Челябинской области
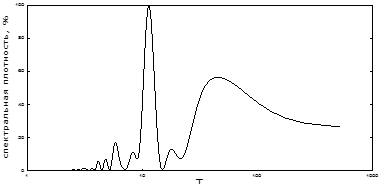
Рис. 2. Спектральный анализ популяционной динамики непарного шелкопряда в Челябинской области
Примерно с таким же периодом на территории Западной Сибири существует ритм суровости зим [6]. В высокой полосе частот мы идентифицировали также особенно много, хотя и небольших по мощности, примерно 2-3-летних циклов (табл. 1). Это самые распространенные практически на всей территории Палеарктики климатические ритмы и ритмы урожайности растений [13]. Таким образом, наиболее мощным ритмом является 10-12-летний. Он вполне может синхронизироваться к обычному здесь циклу летней атмосферной циркуляции и к температурному циклу приблизительно того же периода [9; 19].
Таблица 1. Соотношение величины и мощности периодических составляющих многолетней динамики вспышек массового размножения непарного шелкопряда в Челябинской области
|
Период, год Популяция |
15-50 |
6-12 |
4,1-5,9 |
3-4 |
2,0-2,8 |
|
«Челябинская» географическая популяция непарного шелкопряда |
37.1 25837 |
11.3 54191 |
4.8 4591 |
3.6 1175 |
2.7 611 |
|
17.9 8745
|
8.2 6983 |
4.1 3387 |
3.0 898
|
2.5 461 |
|
|
5. 8 9871 |
2.3 223 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плотн.)
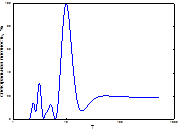
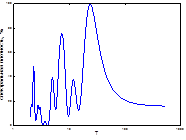
Как известно, периодичность вспышек массового размножения инициируется цикличностью весенне-летних засух [1; 4; 16]. Как показано нами ранее, на Южном Урале они наиболее часто предшествуют вспышкам массового размножения и синхронизированы с продромальной и частью эруптивной фаз популяционной динамики [1; 16]. При этом важное значение имеет характер засух, их интенсивность. Для успешной реализации вспышки массового размножения наиболее эффективными являются полные весенне-летние засухи, не прерываемые периодами с высокими показателями ГТК, особенно майские и июньские [1; 17]. Из этого следует, что полной синхронизации ритмов ГТК с ритмом доминантной составляющей циклов может и не быть. Механизм вспышек массового размножения изучен нами ранее [1; 17; 18]. Для анализа вклада весенне-летних засух в ритмы вспышек массового размножения непарного шелкопряда на Урале и степени их синхронности мы использовали многолетние данные по динамике гидротермических коэффициентов Селянинова. Как показали результаты, ход динамики ГТК в весенне-летние месяцы достаточно сложен и разнообразен (рис. 3). Поэтому для детального анализа взаимосвязи ГТК с циклами вспышек массового размножения непарного шелкопряда были рассчитаны частотные спектры их изменений (рис. 4). Как видно из рис. 4, эти спектры похожи между собой. Различаются они локализацией доминирующего пика, а также количеством и мощностью высокочастотных гармоник. Для уточнения рассчитаем периоды и мощности периодических составляющих (табл. 2).
Как показали результаты спектрального анализа, количество циклов засух имеет частичные отличия от циклов вспышек у этой географической популяции непарного шелкопряда. Это наличие 13-20-летнего цикла ГТК, который проявляется в наибольшей степени в мае (мощность: 3,19) и в значительно меньшей степени в августе и сентябре (мощность, соответственно, 0,39 и 0,43). Все остальные циклы: как низкочастотные (32,5) и (37,5) имеют близкое сходство, так и более короткие циклы практически идентичны. Это 6-10-летний, 4.1- 5,5-летний, 3-4-летний и 2-2,8-летний (табл. 2). Не рассматривая все спектры динамики ГТК, остановимся только на тех, периоды которых близки
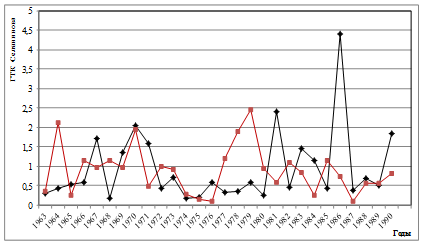
Рис. 3. Многолетняя динамика ГТК Селянинова в весенне-летний период (♦- май, ■- июнь)
 1
1 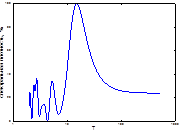 2
2 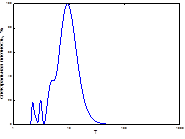 3
3
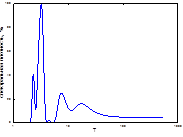 4
4 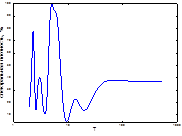 5
5 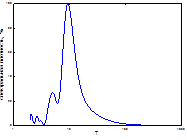 очаг
очаг
Рис. 4. Спектральный анализ динамики колебаний ГТК в Челябинской обл.
Примечание: слева-направо: апрель, май, июнь, август, сентябрь, спектр динамики вспышек.
по значению гармоническим составляющим хода динамики численности непарного шелкопряда. Можно заметить, что наибольшее совпадение у динамики его популяции с 6-10-летними циклами весенне-летних засух. Это также цикл с наибольшей мощностью в апреле (24,78), июне (3,5) и слабой в августе (0,61) (табл. 2). Обращает внимание полное отсутствие в 6-10-летней цикличности ГТК майских ритмов. Но майские засухи имеют особенно важное значение для роста и развития гусениц непарного шелкопряда, отродившихся из кладок [16].
Таблица 2. Соотношение величины и мощности периодических составляющих многолетней динамики ГТК Селянинова в Челябинской области
|
Период, год
ГТК |
30-70 |
13-20 |
6-10 |
4,1-5,5 |
3-4 |
2,0-2,8 |
|
Апрель |
32.50 2.86 |
|
9.81 24.78 |
5.00 3.19 |
3.02 6.92 |
2.25 2.40 |
|
4.00 1.49 |
||||||
|
Май |
|
14.86 3.19 |
|
5.25 1.10 |
3.71 0.48 |
2.71 1.16 |
|
2.41 0.98 |
||||||
|
2.01 0.80 |
||||||
|
Июнь |
|
|
9.29 3.50 |
5.00 1.29 |
3.06 0.72 |
2.20 0.64 |
|
Август |
|
17.33 0.39 |
7.43 0.61 |
4.41 0.06 |
3.21 2.42 |
2.29 0.99 |
|
Сентябрь |
65.00 0.73 |
13.68 0.43 |
|
5.05 1.92 |
3.02 0.77 |
2.29 1.49 |
|
Очаг |
|
|
9.29 2008 |
4.91 543
|
3.02 77
|
2.57 154 |
|
2.01 189 |
В этот период они наиболее чувствительны к качеству кормового субстрата и температуре. Майский ритм с высокой мощностью (3,29) присутствует в составе 13-20-летнего цикла колебаний ГТК (табл. 2). Кроме того, он имеется у 2, 3 и 4-летнего цикла. На основании этого можно предполагать, что в годы, когда этот 6-10-летний ритм совпадает с другими ритмами, площадь очагов может заметно возрастать. Но, учитывая мощность майского цикла ГТК у 13-20-летнего ритма ГТК, их наложение сопровождается возникновением особенно крупных по площади вспышек массового размножения непарного шелкопряда (рис. 1). Остальные, более короткие ритмы колебаний ГТК, вероятно, способствуют переходу популяций этого фитофага на более короткие ритмы популяционной динамики, которые наблюдаются у других географических популяций в условиях большей частоты возникновения весенне-летних засух. В целом изучение особенностей спектральных параметров вспышек массового размножения непарного шелкопряда позволило установить, что они имеют скрытые периодические составляющие. Эти циклы хорошо проявляются на спектре.
Спектральный анализ многолетней динамики вспышек массового размножения непарного шелкопряда в Свердловской обл.
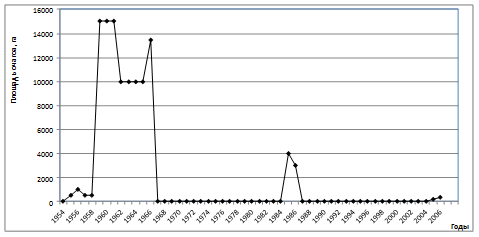
Рис. 5. Многолетняя динамика вспышек массового размножения непарного шелкопряда в Свердловской области
Анализ многолетней динамики вспышек массового размножения непарного шелкопряда в Свердловской обл. показал, что за весь срок наблюдений было три вспышки массового размножения. Первая, наиболее продолжительная, продолжалась восемь лет (рис. 5). Она имела два максимума, с незначительным понижением численности между ними. Она же была и самой крупной по площади за 52 года (рис. 5). Интервал между первой и второй вспышками составлял 17 лет.
Спектральный анализ динамики вспышек этой популяции показал, что сложная многолетняя динамика численности этого вида представляет собой суперпозицию 8 гармоник, одна из них, примерно 40-45-летняя, - самая мощная (рис. 6). Ее присутствие мы не могли предполагать при анализе графика многолетней динамики вспышек массового размножения этого фитофага. Остальные циклы на спектре хотя и проявлены, но заметно уступают в мощности самой низкочастотной гармонике. Среди них заметной мощностью обладает более чем 10-летняя. Имеются и проявленные гармонические составляющие в диапазоне высоких частот. Последние идентичны ритмам популяционной динамики непарного шелкопряда в Челябинской обл. (табл. 1, 3). Значения периодов и мощностей этих гармонических составляющих приведены в табл. 3. Самый мощный ритм, близкий к 45-летнему, вполне может синхронизироваться к обычному циклу чередования прохладно-влажных и тепло-сухих периодов в интервале 35-45 лет по А.В. Шнитникову [32], или к внутривековой изменчивости климата 35-45лет [10; 20; 21]. Существуют здесь и гидроклиматические циклы, а также георитмы Земли, имеющие периодичность 7-12 лет [21; 33]. С их помощью популяция может адаптироваться к некоторым, близким по периоду, ритмам внешней среды. Близкий к 5-летнему цикл характерен для июльско-августовских осадков [9] и может «затягиваться» соответствующим по величине георитмом Земли [33]. Примерно с таким же периодом на территории Западной Сибири есть ритм суровости зим [6]. В полосе высоких частот у динамики непарного шелкопряда особенно много небольших по мощности примерно 2-4-летних циклов. К ним принадлежат самые распространенные практически на всей территории Палеарктики климатические ритмы и ритмы урожайности растений [13].
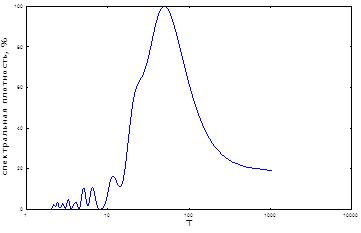
Рис. 6. Спектральный анализ динамики численности непарного шелкопряда в Свердловской области
Таблица 3. Соотношение величины и мощности периодических составляющих многолетней динамики очагов непарного шелкопряда в Свердловской области
|
Период, год очаг |
40-60 |
10-15 |
5-7 |
3,5-4,5 |
2-3 |
|
«Свердловская» географическая популяция непарного шелкопряда |
43.33 185
|
11.69 30
|
5.05 19 |
4.05 6 |
2.41 6 |
|
6.46 19
|
3.26 9 |
2.79 5 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плотн.) (1988).
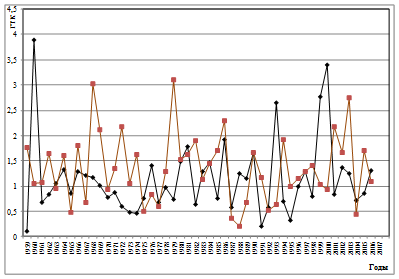
Рис. 7. Многолетняя динамика ГТК Селянинова в Свердловской обл. (♦-май, ■-июнь).
Мы проанализировали ГТК мая-августа (рис. 4), включив в эту иллюстрацию и спектр периодов непарного шелкопряда. Сравнение их дало возможность изучить степень синхронизации спектров популяционной динамики непарного шелкопряда со спектральными параметрами гидротермических коэффициентов Селянинова. Можно заметить, что спектры динамики ГТК похожи между собой даже в локализации самого низкочастного пика (табл. 2, 4). Число высокочастотных гармоник на всех спектрах велико, а у отдельных они доминируют по мощности (рис. 8). Кроме этого, проведен расчет периодов и мощности периодических составляющих (табл. 4). Все циклы по периодичности примерно совпадали с периодами, характерными для климатических условий, посчитанных нами для Челябинской области (см. выше). Самый мощный пик также имел отличия по периоду. В условиях Челябинской обл. он составлял 6-10 лет, а Свердловской - 10-15 (табл. 2, 4). Отличался также и более длинный ритм. В условиях Свердловской обл. он составлял 20-30 лет, а Челябинской – 13-20. Как и у предыдущего варианта, в Свердловской обл. в спектре ритмов колебаний ГТК отсутствовал майский цикл. Следовательно, в Южном Зауралье основные ритмы ГТК были несколько короче.
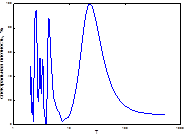 1
1 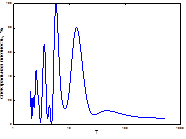 2
2 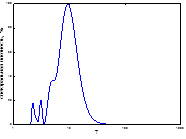 3
3
 4
4 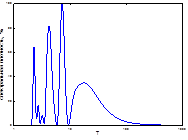 5
5 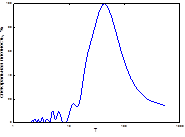 очаг
очаг
Рис. 8. Спектры периодов в динамике ГТК (1 май, 2 июнь, 3 июль, 4 август, спектр очага).
Заметим, что для климатических условий Свердловской обл. характерна близкая к 20-летней цикличность колебаний ГТК (май, июль). Но идентичная цикличность отсутствует в составе спектра популяционной динамики «Свердловской» географической популяции непарного шелкопряда. Следовательно, это может быть одной из основных причин, по которым 20-летний цикл популяционной динамики этого фитофага в этих климатических условиях не может быть реализован. Цикличность в средних и высоких частотах дает возможность синхронизации динамики численности с ритмами ГТК практически все летние месяцы.
Спектральный анализ динамики вспышек массового размножения «Оренбургской» географической популяции непарного шелкопряда
Как известно [28], за весь срок наблюдений были отмечены три вспышки массового размножения непарного шелкопряда. При этом интенсивность каждой последующей нарастала. Особенно крупной по площади была последняя вспышка, вдвое превзошедшая предыдущую (рис. 9). Интервалы между вспышками составляли, соответственно, 6; 6 и 8 лет (в среднем 6,7 года). Продолжительность каждой вспышки массового размножения составляла 4-5 лет. Как показали результаты спектрального анализа, в составе спектра оказалось всего три периодических составляющих. Самая мощная из них - в средних частотах - близкая к 8-летней, вторая – 3,5 года, третья – 2,32 года (табл. 5). В направлении к высоким частотам периоды гармоник постепенно убывали по мощности (рис. 10). Динамика вспышек непарного шелкопряда в Оренбургской области имеет несколько различных по мощности скрытых периодичностей (рис. 10). С их помощью популяция способна адаптироваться к некоторым, близким по периоду, ритмам внешней среды. Мы рассчитали значения периодов и мощностей этих гармонических составляющих (табл. 5). Как показали результаты, самый мощный ритм был близким к 7,5-летнему (табл. 5). Он присущ многим атмосферным процессам. Следующая периодичность в три раза менее мощная – близкая к 3,5-летней. Она почти кратная первому циклу. Поэтому вполне вероятно, что она участвует в климатических ритмах и ритмах урожайности растений [13].
Таблица 4. Соотношение величины и мощности периодических составляющих многолетней динамики ГТК Селянинова в Свердловской области
|
Период, год ГТК |
40-60 |
20-30 |
10-15 |
5-7 |
3,5-4,5 |
2-3 |
|
Май |
|
23.64 1.06
|
|
|
4.26 0.94 |
2.51 1.01 |
|
2.96 0.59 |
2.1 0.20 |
|||||
|
Июнь |
47.27 0.41 |
|
13.33 2.83 |
5.71 3.51 |
3.54 2.34 |
2.52 1.58 |
|
Июль |
|
23.64 4.77
|
11.82 1.81
|
7.32 3.60
|
4.95 1.88 2.96 |
2.72 0.80 |
|
0.74
|
2.27 2.34 |
|||||
|
Август |
|
|
17.33 1.32
|
7.22 3.76
|
4.19 3.09 |
2.27 2.44 |
|
3.17 0.31 |
2.69 0.63 |
|||||
|
Очаг |
43.33 217753760.71
|
|
12.09 35334099.34
|
5.05 21694356.25 |
4.03 6223056.86 |
2.40 7580357.60 |
|
6.50 20917744.48 |
3.27 9578600.02 |
2.78 5441338.19 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектральной плотности)
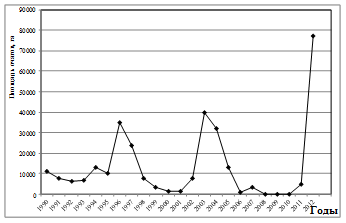
Рис. 9. Многолетняя динамика вспышек массового размножения непарного шелкопряда в Оренбургской области
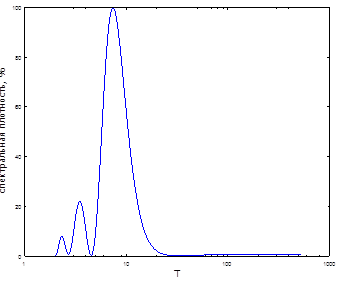
Рис. 10. Спектр ритмов динамики численности непарного шелкопряда в Оренбургской области
Наименее мощная периодичность - близкая к 2,3-летней. Можно предполагать, что она способна синхронизироваться с цикличностью, близкой к 3,5-летней. В результате это индуцирует реализацию самой мощной гармоники на этом спектре. Продолжительность наблюдений в Оренбургской области была относительно небольшой (23 года). Гармонический анализ позволил выделить всего три периодичности и не обнаружил более низкочастотных, хотя этот метод расчета обычно фиксирует наличие гармоники, соизмеримой с длиной временного ряда. Вероятно, на территории Оренбургской обл. у локальной популяции непарного шелкопряда такая цикличность отсутствует.
Таблица 5. Соотношение величины и мощности периодических составляющих многолетней динамики очагов непарного шелкопряда в Оренбургской области
|
Период, год Очаг |
5 – 9 |
4,9 – 3,1 |
2,0 – 3,0 |
|||
|
период |
мощность |
период |
мощность |
период |
мощность |
|
|
Непарный шелкопряд |
7.43 |
1407 |
3.49 |
310 |
2.32 |
112 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плотн.)
Спектральный анализ многолетней динамики вспышек массового размножения непарного шелкопряда на Зилаирском плато (Башкортостан)
Чтобы проверить наличие возможных скрытых периодических составляющих в динамике вспышек массового размножения непарного шелкопряда, был проведен спектральный анализ популяционной динамики этого фитофага на Зилаирском плато (Башкортостан) со значительно более длинным временным рядом (208 лет) [22]. Среднее значение интервала - 9,4 года. По мнению автора, такова основная цикличность вспышек размножений этого вида в условиях Зилаирского плато [22].
Мы провели спектральный анализ ритмов динамики численности «Башкирской» географической популяции непарного шелкопряда. Как видно из рис. 11, на спектре оказалось много гармонических составляющих, большинство из которых имели пики правильной экспоненциальной формы с узкими основаниями. В области низких частот (100-200 лет) основания их увеличивалось, пики сглаживались (рис. 11). Наибольшей мощностью обладали две периодичности. Одна из них чуть менее 10 лет, видимо, та, что С.Е. Кучеров получил как среднеарифметическую, вторая же несколько больше 10-летней (рис. 3). Как известно, все биологические колебания устроены так, что их частота (период), амплитуда и фаза всегда «колеблются». Такая особенность устройства облегчает их согласование с ритмикой внешней среды [25]. К тому же в любой полосе частот у популяции обычно бывает несколько периодических составляющих, близких по величине. Рассчитанный нами спектр ритмов очага непарного шелкопряда из Зилаирского плато имел именно такую спектральную характеристику. В каждой полосе частот на нем находится по целому «семейству» гармоник (рис. 11). Расчет характеристик этих периодических составляющих показал, что в каждом таком «семействе» по несколько близких ритмов, но они могут различаться по мощности (табл. 6). Наиболее мощные циклы вспышек массового размножения – близкие к 9-10-летним. Очень близка по мощности к ним и около 7-летняя периодичность (табл. 6). В каждой полосе высоких частот обнаруживаются по 2-4 гармонических составляющих, сходных по периоду. С их помощью популяция непарного шелкопряда может легко «подстраиваться» к нужным ей колебаниям среды. Несколько ритмов облегчают такую подстройку, потому что чем ближе период к таковому у внешнего датчика времени, тем легче происходит синхронизация к нему.
Низкочастотные ритмы вспышек массового размножения у этой популяции включают относительно мощный, близкий к 20-летнему, цикл (табл. 6). Он близок к циклу осадков [9] и георитмам Земли [26]. Заметный по мощности, близкий к 30-летнему, цикл соответствует многократно описанному климатическому (брикнеровскому) циклу. Значительная длина временного ряда позволила выявить еще один из низкочастотных ритмов вспышек массового размножения этой популяции. Он плохо проявлен на спектре, и пришлось изменить окно при расчетах для повышения детализации. При этом появляется опасность появления «ложных» пиков на спектре. При таком расчете мы уточнили характеристики этой гармоники. Ее период оказался примерно 130-летний, мощность же не изменилась. Все прочие гармонические составляющие остались в пределах своих характеристик. Такая периодичность может существовать и поддерживаться близкими по величине
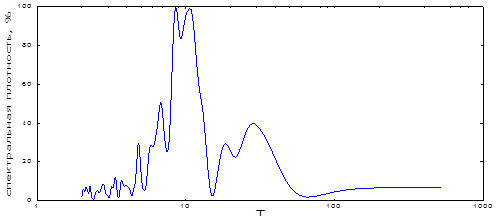
Рис. 11. Спектральный анализ динамики численности непарного шелкопряда в Башкортостане.
Таблица 6. Соотношение величины и мощности периодических составляющих многолетней динамики очагов непарного шелкопряда в Башкортостане
|
Период, год
Популяция |
100-300 |
20-30 |
12-15 |
10-11 |
5-9 |
4,9-3,1 |
2-3 |
|
«Башкирская» популяция непарного шелкопряда (Зилаирское плато) |
260.00 0.17
|
28.89 1.01
|
18.57 0.75 |
10.61 2.53
|
8.53 2.55 |
4.82 0.75 |
2.78 0.22 |
|
6.75 1.29 |
3.69 0.26 |
2.55 0.13 |
|||||
|
2.27 0.20 |
|||||||
|
2.13 0.18 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плот.)
брикнеровскими и солнечными ритмами, у них отмечены вековые колебания [10; 35] но для того, чтобы утверждать это, нужно получить его в виде хорошо выраженного на спектре пика с узким основанием. Для этого ряд должен быть длиннее в несколько раз. Можно заметить, что многолетняя динамика численности непарного шелкопряда имеет характерные внутренние цикличности, с помощью которых она может совершать «подстройку» к колебаниям внешних факторов. В различных географических популяциях этого фитофага используются разные периодические составляющие из имеющегося спектра, и это можно видеть по изменениям их мощностей на разных спектрах.
Спектральный анализ многолетней динамики вспышек массового размножения «европейской» популяции непарного шелкопряда
Е.Н. Белецким [2] приведен почти 150-летний ряд динамики вспышек популяции непарного шелкопряда в Европе. Мы, применив спектральный анализ, изучили спектр ритмов этой популяции. Как показали результаты, наибольшей мощностью обладает близкая к 9-летней периодичность, значительная по мощности – близкая к 12-летней периодичность, несколько ниже – близкая к 5-летней (табл. 7). Кроме того, имеется несколько гармонических составляющих незначительной мощности в области высоких частот (2-3-летние). Возможно, этот набор периодов видоспецифичен для непарного шелкопряда. Имеется на спектре и примерно 40-летний ритм численности, так что, возможно, в этом спектре проявилась действительно имеющая место внутривековая (брикнеровская) климатическая ритмичность в динамике очагов.
Обсуждение
Нами проведен спектральный анализ популяционной динамики 5 географических популяций непарного шелкопряда. Исследования показали, что в трех из них («Европейской», «Оренбургской» и «Башкирской») по мощности преобладал одинаковый цикл (7,5-8,8 года). В двух остальных он отсутствовал и заменялся на 11,5-летний. В популяции на северной границе очагов доминировал 43-летний цикл, который выявлен у популяций в Европе и Челябинской обл. Все высокочастотные циклы (2-3-летний и 3-5-летний) у всех изученных популяций были идентичны. Что касается наиболее низкочастотного цикла (260 лет), то более короткие временные ряды не дают возможности выявить этот цикл. В популяциях, локализованных в более южных широтах, доминантные по мощности циклы вспышек массового размножения были короче, чем в более северных популяциях. В наименее благоприятных климатических условиях преобладал еще более низкочастотный, 43-летний цикл. Тем не менее у всех изученных популяций непарного шелкопряда имеются идентичные циклы популяционной динамики.
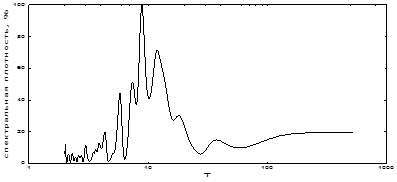
Рис. 12. Спектральный анализ динамики численности «Европейской» популяции непарного шелкопряда
Таблица 7. Соотношение величины и мощности периодических составляющих многолетней динамики очагов «Европейской» популяции непарного шелкопряда
|
Период, год
Популяция |
35-45 |
16-20 |
12-15 |
8-10 |
4-6 |
2-3 |
|
Непарный шелкопряд, «Европейская» популяция |
37.14 0.44
|
17.93 0.90
|
12.38 2.01
|
8.81 2.95
|
5.78 1.32 |
2.99 0.35 |
|
4.33 0.58
|
2.60 0.15 |
|||||
|
2.47 0.11 |
||||||
|
2.14 0.18 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плотн.)
Исходя из полученных результатов можно предполагать, что набор популяционных циклов у насекомых-фитофагов с высоким биотическим потенциалом очень консервативен и в процессе эволюции был синхронизирован с частью циклов колебаний ГТК. Это вполне обоснованно с точки зрения возможности реализации жизненной стратегии этих видов, направленной на сохранение существующей стратегии освоения жизненного пространства и своей экологической роли в лесных фитоценозах: сохранении устойчивых ценопопуляций древесных растений и элиминации ослабленных древостоев за счет дефолиации крон.
Исходя из этого можно предполагать, что у всех изученных нами географических популяций непарного шелкопряда, обитающих в различных климатических условиях, очевидно, имеется полный набор всех типов низкочастотных циклов популяционной динамики, который не выявляется вследствие коротких временных рядов у некоторых популяций. По нашему мнению, именно полный набор циклов детерминирует наибольшие возможности у вида адаптироваться к динамике колебаний климатических факторов, чтобы реализовать жизненную стратегию. Но анализ климатических условий, предшествующих возникновению вспышек численности и синхронизированных с ними, однозначно показывает, что наиболее часто в этот период преобладают майско-июльские сочетания низких ГТК (Колтунов, 2006, Колтунов и др., 2010), которые не свойственны доминантным циклам популяционной динамики и колебаний ГТК. Поэтому мы предполагаем, что такое сочетание ГТК могло возникнуть в основном в результате наложения ряда циклов и эволюционной адаптации к ним циклов популяционной динамики непарного шелкопряда. Как показали результаты, с помощью спектрального анализа обнаруживаются скрытые цикличности.
Рецензенты:
Усольцев Владимир Андреевич, доктор сельскохозяйственных наук, профессор, профессор Уральского государственного лесотехнического университета, г. Екатеринбург.
Менщиков Сергей Леонидович, доктор сельскохозяйственных наук, заведующий отделом лесоведения ФГБУН «Ботанический сад Российской академии наук», г. Екатеринбург.
Библиографическая ссылка
Колтунов Е.В., Ердаков Л.Н. СПЕКТРАЛЬНЫЙ АНАЛИЗ МНОГОЛЕТНЕЙ ДИНАМИКИ ВСПЫШЕК МАССОВОГО РАЗМНОЖЕНИЯ НЕПАРНОГО ШЕЛКОПРЯДА (LYMANTRIA DISPAR L. ) НА УРАЛЕ // Современные проблемы науки и образования. 2013. № 2. ;URL: https://science-education.ru/ru/article/view?id=8625 (дата обращения: 01.07.2026).