


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
MODELLING OF ADAPTIVE SELF-ORGANIZING OF ECOSYSTEMS

Введение
Среди наиболее актуальных проблем современности выделяется проблема выяснения причин и пределов устойчивости сложных живых систем, состоящих из огромного числа компонентов. К таким системам относятся организмы и биосфера с ее экосистемами. В условиях планетарного экологического кризиса от нашего умения понимать и предсказывать поведение экосистем и самой биосферы зависит выживание человечества.
Многие отечественные и зарубежные ученые изучают эту проблему с разных сторон на основе различных представлений. В частности, делаются все более многочисленные попытки ухода от классических консервативных (механистических) моделей, не учитывающих фундаментальные свойства живых систем. Можно выделить два основных направления исследований.
Первое заключается в поиске простых представлений, позволяющих отразить наиболее значимые для авторов моделей свойства живых систем и попытаться на их основе (с привлечением математической теории катастроф и др.) спрогнозировать ключевые процессы в экосистемах [2, 11].
Второе, наоборот, акцентируется на сложности как важнейшей характеристике живого и пытается предложить способы построения моделей, учитывающие эту сложность и вытекающие из нее базовые свойства экосистем [3].
Взаимная критика сводится к следующему: 1) сторонники первого направления апеллируют к необозримости и трудностям анализа сложных моделей; 2) представители второго направления указывают на чрезмерное упрощение и схематичность простых моделей, скрывающие важные свойства природных систем, что ограничивает возможности их понимания и прогноза. В целом же, два этих направления дополняют друг друга, давая различные перспективы (масштабы) граней отображаемой реальности.
Вместе с тем общей трудностью построения классических и неоклассических моделей является необходимость детального выстраивания их вручную, требующая больших усилий и времени и не позволяющая выполнять быстрые мобильные перестройки при сильном изменении условий или для отличающихся задач.
Эти трудности приобретают не только технический, но и фундаментальный характер при переоценке роли соотношения структуры и функции в природных системах в свете современных знаний. Так, в отличие от классических представлений, структура связей и вид функций в экосистемах (и других природных системах) не статичны и могут изменяться при изменении внешних условий (включая антропогенные воздействия) не только благодаря образованию новых видов, но и в силу адаптации к изменению особенностей взаимодействий между видами.
Результатом таких изменений является динамическая перестройка аттрактивого ландшафта системы, на котором базируется многообразие ее поведения[1]. Множество квазистационарных состояний (аттракторов, инвариантных многообразий) формируют множественную устойчивость системы в ответ на изменение внешних условий (инвариантный портрет системы).
Такая организация позволяет избежать фундаментальных проблем, проявляющихся в классической методологии моделирования сложных природных систем, известных как "проклятье размерности" и "проклятье неустойчивости". Само их наличие указывает на глубокое противоречие в понимании и теоретическом описании количественных законов функционирования природных систем с наблюдаемой реальностью. В частности, известно, что в подавляющем большинстве случаев устойчивость экосистем растет с ростом их сложности и видового разнообразия [13, 10], а также при учете в моделях адаптивного метаболизма [14].
В предыдущих работах [9, 6, 7] изложена новая высоко эффективная методология построения моделей природных систем, основывающаяся на Концепции Адаптивной Самоорганизации сложных природных систем (КАС), предложенной Ю. П. Ланкиным, которая базируется на отражении их фундаментальных свойств: сетевая структура, адаптивность, аттрактивность, фрактальность. В публикации [7] представлена модель адаптивной самоорганизации на основе уравнений Ферхюльста для описания динамики лесообразования множества взаимодействующих видов.
Данная работа демонстрирует возможность перехода (связующего "моста") между классическими и новыми методами моделирования [8]. Ранее, в книге [12] была предложена классическая модель, не учитывающая возможность адаптации организмов, населяющих экосистему, что на практике должно приводить к изменению системных свойств. В данной работе эта модель дополнена механизмом адаптивной обратной связи, который корректирует структуру связей системы, пытаясь возвратить ее в стационарное состояние. И тем самым обеспечивает динамическую реакцию на нарушение устойчивости экосистемы. Имитация механизмов обратной связи в экосистеме выполняется с помощью нейронных сетей, предложенных в [4, 5].
Модель экосистемы
Модель экосистемы, представляющая собой систему дифференциальных уравнений первого порядка, предложена Н. С. Печуркиным в книге [12]. Система дифференциальных уравнений, соответствующая замкнутой экосистеме, содержит следующие обозначения: S, P, C, D - концентрации биогенных элементов, продуцентов, хищников и детрита, соответственно; μ2, μ3, μi - удельные скорости роста продуцентов и хищников 1-го и i-го родов (i=3...n); F - скорость поступления лимитирующего вещества; S0 - начальная концентрация биогенных элементов; Y2, Y3, Yi - экономический коэффициент роста продуцентов, хищников 1-го рода и хищников i-го рода; ε2, εi - удельные скорости отмирания продуцентов и хищников; εd - скорость разложения детрита; q - коэффициент возврата биогенных элементов в экосистему. Модель имеет следующий вид:
 (1)
(1)
Для исследования эффективности предложенных адаптивных алгоритмов была использована версия данной модели из работы [1]. Эта усовершенствованная версия интересна тем, что содержит замкнутый трофический цикл, благодаря добавлению уравнения для редуцентов X5, и каналы обмена с внешней средой по каждому компоненту (рис.1).
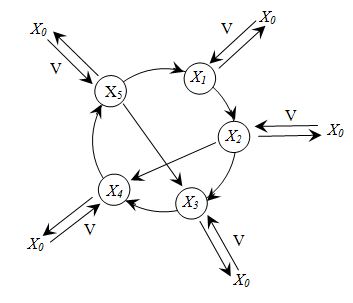
Рис.1. Схема экосистемы с циклом по лимитирующему элементу и множественными каналами обмена с внешней средой. Обозначения: X1 - субстрат; X2 - продуцент; X3 - консумент; X4 - детрит; X5 - редуцент [5]
Измененная модель (1) описывается в [5] следующей системой уравнений:
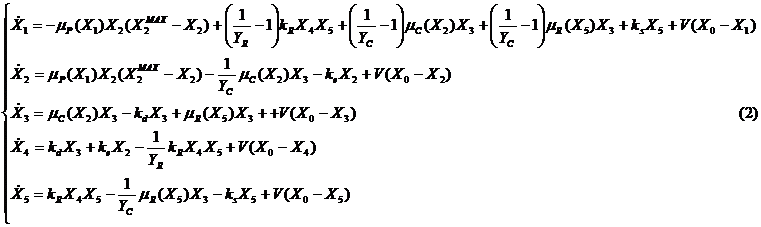
где: ![]() ,
, ![]() ,
, ![]() - удельные скорости роста продуцентов, консументов, редуцентов, соответственно; kR, ke - удельные скорости отмирания редуцентов и продуцентов; ks , kd - скорости потребления субстрата и разложения детрита; YR, YC - экономические коэффициенты роста редуцентов и консументов; VP, VC, VR - интенсивности взаимодействия продуцентов, консументов и редуцентов с внешней средой; KP, KC, KR - значения коэффициентов чувствительности продуцентов, консументов и редуцентов; V - скорость поступления в систему соответствующего компонента; X0, - концентрация вещества или численность организмов данного вида; X2MAX - максимально-возможная численность продуцентов.
- удельные скорости роста продуцентов, консументов, редуцентов, соответственно; kR, ke - удельные скорости отмирания редуцентов и продуцентов; ks , kd - скорости потребления субстрата и разложения детрита; YR, YC - экономические коэффициенты роста редуцентов и консументов; VP, VC, VR - интенсивности взаимодействия продуцентов, консументов и редуцентов с внешней средой; KP, KC, KR - значения коэффициентов чувствительности продуцентов, консументов и редуцентов; V - скорость поступления в систему соответствующего компонента; X0, - концентрация вещества или численность организмов данного вида; X2MAX - максимально-возможная численность продуцентов.
Значения параметров: kR = 0.5, ke = 0.02, ks = 0.05, kd = 0.1, YR = 0.2, YC = 0.5, VP = 5, VC = 0.2, VR = 0.1, KP = 0.4, KC = 0.4, KR = 0.5, V = 1, X0 = 1, X2MAX = 1.
Динамика взаимодействия видов в модели экосистемы для приведенных значений параметров отображена на рис.2 в программе сеточного решения и анализа систем дифференциальных уравнений NetworkDecisions, разработанной Ю. П. Ланкиным.
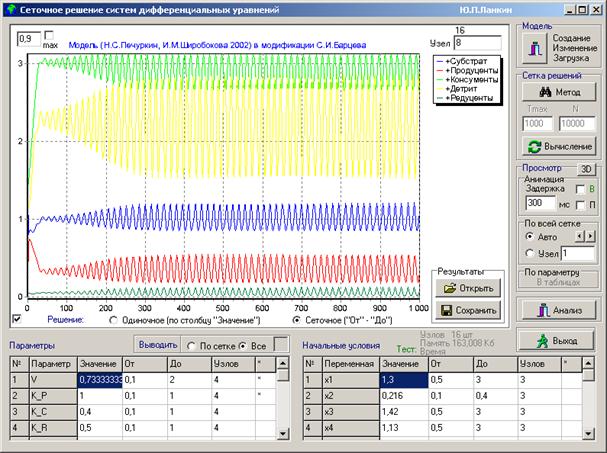
Рис.2. Колебательный режим, в который классическая (консервативная) модель экосистемы (2) переходит после потери устойчивости
Моделирование адаптивной самоорганизации экосистемы с помощью нейронной сети
Проведение исследований по численному моделированию адаптивной самоорганизации модели экосистемы к стационарным состояниям было выполнено на компьютерной программе, разработанной Ю. П. Ланкиным. Внешний вид программы приведен на рис.3.
Программа содержала два взаимодействующих модуля:
1) модуль решения системы дифференциальных уравнений модели экосистемы методом Рунге - Кутта с постоянным шагом;
2) модуль нейронной сети, имитирующий адаптивную обратную связь в экосистеме и выполняющий динамическую коррекцию параметров (структуры) модели непосредственно в процессе ее функционирования ("жизнедеятельности").
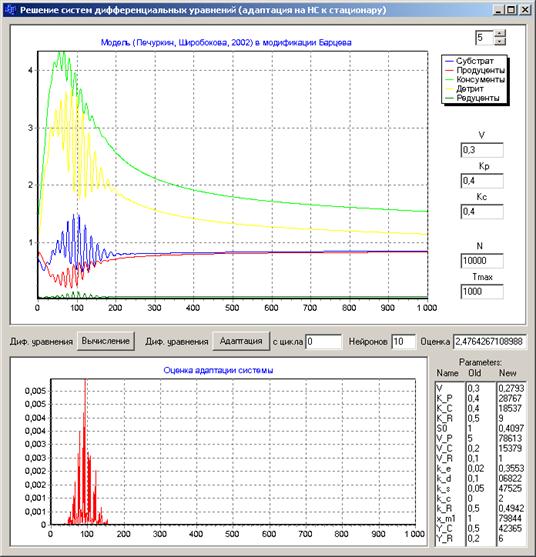
Рис.3. Программа моделирования динамического процесса восстановления стационарного состояния модели экосистемы (после потери ей устойчивости) с помощью предложенной нейронной сети (верхний график). На нижнем графике показана кривая оценки величины отклонения модели экосистемы от стационарного состояния
Исходное поведение модели экосистемы (2) приведено на рисунке рис.2. Система теряет устойчивость (неустойчивый узел) и переходит в режим колебаний. В программе рис.3 этот режим воспроизводится кнопкой "Вычисление". Кнопкой "Адаптация" запускается режим адаптивной самоорганизации модели (системы дифференциальных уравнений) к стационарному состоянию, изображенный на этом рисунке. В правой нижней части формы программы рис.3 отображены параметры экологической модели (2): столбец "Name" - обозначение параметров в программе, столбец "Old" - начальные значения параметров, столбец "New" - новые значения параметров, подбираемых нейросетью и выводимых при каждом их изменении в процессе работы программы. Расшифровка параметров приведена выше при описании модели (2).
На рис.4 представлен другой вариант динамики адаптивной самоорганизации модели экосистемы нейронной сетью. При запуске системы дифференциальных уравнений из одних и тех же начальных условий различие достигается генерацией случайных начальных значений весов связей нейронной сети. Таким способом удается отобразить разнообразие поведения живых организмов (видов) в экосистеме во внешне сходных ситуациях. Легко видеть, что разнообразие в поведении видов не сказывается на возможности достижения экосистемой стационарного состояния. Различие заключается лишь в формировании разных аттракторов (инвариантных многообразий).
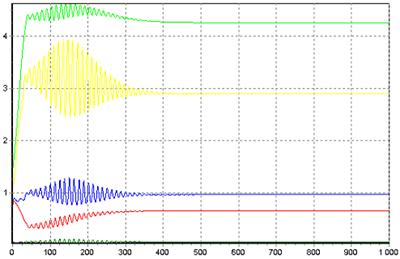
Рис.4. Адаптивная самоорганизация структуры модели экосистемы, возвращающая ее из режима колебаний (рис.2) к стационарному состоянию (формирование аттрактора)
Нейронные сети реализовали адаптивную самоорганизацию модели экосистемы путем динамического построения отображений из 5-мерного пространства переменных модели экосистемы (системы дифференциальных уравнений) в 16-мерное пространство ее параметров.
Специфические особенности процесса моделирования связаны с нестационарностью процесса решения задачи из-за постоянно изменяющихся параметров (коэффициентов) системы дифференциальных уравнений (модели), различие иерархий времен для переменных и параметров модели и необходимость совместного решения систем дифференциальных (модель) и алгебраических (нейросеть) уравнений.
Благодаря нелинейности нейросеть функционировала только по точкам графика (один раз в 10 вычислений производных модели), что на порядок сокращало затраты времени вычислений относительно модели.
Нейросеть моделировала адаптивные процессы во взаимосвязанной сети организмов (видов) при самоорганизации экосистемы к стационарному состоянию и функционировала в замкнутом цикле следующим образом:
- На каждом шаге решения на вход нейросети подавались пять значений переменных системы дифференциальных уравнений;
- Система алгебраических уравнений формальных нейронов в нейронной сети функционировала, генерируя решения нейронов в виде αik+1;
- Целевая функция H оценивала величину отклонения модели от стационарного состояния;
- Вычислялись новые значения величин связей между элементами сети xij, аналогичных по смыслу силе связей между организмами экосистемы (или, в усредненном виде, между видами);
- Корректировались значения параметров модели (системы дифференциальных уравнений) pik+1 = pik + ci * αik+1, где pi - параметры модели.
Затем цикл повторялся вплоть до достижения моделью стационарного состояния (аттрактора), выражающегося в стабилизации численностей моделируемых видов и прекращении изменений параметров.
Устойчивость моделей с адаптивной самоорганизацией
Адаптивная самоорганизация системы к стационарным состояниям непосредственно в процессе функционирования системы приводит к перестройке ее структуры, а следовательно, и к динамическому изменению структурно-функциональных соответствий.
Одной из важных составляющих исследования поведения моделей является оценка их устойчивости. Обычно для этого используют метод оценки по Ляпунову. Однако в рассматриваемом способе моделирования изменяется сам смысл понятия устойчивость. Попытка вывести такую систему из равновесия активизирует механизмы обратной связи между видами (живыми организмами) в экосистеме, пытающимися вернуть систему в стационарное состояние. Если вернуть ее не удается, то формируется новое стационарное состояние. Число таких состояний (аттракторов) зависит от сложности системы (для модели - числа уравнений и коэффициентов) и с усложнением растет быстрее, чем экспоненциально [1]. Отображение этого свойства лавинообразного увеличения информационной емкости системы (мощности множества инвариантных многообразий) с ростом ее сложности классическими методами практически невозможно и опирается на методы КАС [9, 6, 7].
В отличие от классических консервативных моделей, эти процессы в точности соответствуют процессам, происходящим в экосистемах и в живых организмах.
Заключение
В работе предложен и реализован метод разрешения фундаментальной проблемы "проклятья неустойчивости" сложных моделей природных систем. Эта проблема возникает, наряду с проблемой "проклятья размерности", при попытке достаточно детального описания поведения и структуры природных систем, особенно живых (эко и биосистем). Метод позволяет отразить нелинейные и трудно предсказуемые системные свойства, не улавливаемые в простых моделях. Данная работа демонстрирует возможность построения переходных методов, связывающих сетевые модели адаптивной самоорганизации сложных систем, предложенные ранее [9, 6, 7], с классическими консервативными моделями. Такая возможность появляется благодаря моделированию адаптивных обратных связей в эко и биосистемах с помощью искусственных динамических нейронных сетей, предложенных авторами. Эти обратные связи позволяют вести динамическую перестройку структуры системы в сторону формирования стационарных состояний (аттракторов, инвариантных многообразий), обеспечивающих множественную устойчивость системы с ростом ее сложности. В отличие от традиционных моделей, такая методология соответствует известным фактам о росте устойчивости экологических систем с ростом их сложности и видового разнообразия.
Рецензенты:
- Мысливец Симона Глебовна, доктор физ.-мат. наук, профессор, заведующий кафедрой высшей математики 1, Сибирский федеральный университет, г. Красноярск.
- Сомова Лидия Александровна, доктор биологических наук, ведущий научный сотрудник Института биофизики СО РАН, г. Красноярск.
[1] В качестве наглядной феноменологической аналогии можно привести изменение динамики системы приповерхностных воздушных потоков (на суше) или водных течений (в океане), возникающих при изменении структуры ландшафта под влиянием геологических процессов.
Библиографическая ссылка
Ланкин Ю.П., Басканова Т.Ф., Печуркин Н.С. МОДЕЛИРОВАНИЕ АДАПТИВНОЙ САМООРГАНИЗАЦИИ ЭКОСИСТЕМ // Современные проблемы науки и образования. 2012. № 5. ;URL: https://science-education.ru/en/article/view?id=6735 (дата обращения: 15.07.2026).