


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
CONCEPTION OF DEVELOPMENT LENS IN THE HUMEN EYES

Изучены 171 глаз эмбрионов и плодов человека. Использованы классические гистологические методы исследования с окрашиванием г/э; Victoriablue и импрегнация серебром, Ironhematoxilin, NADFH-диафоразы, а также иммуногистохимические методы на выявление CD4, CD8, CD 68, CD163, CD 204, TUNEL-метод на выявление апоптозирующих клеток, Ki67 для выявления пролиферативной активности капсулярных клеток. Анализ материала проведён с помощью микроскопа Olympus-Bx51 и цифровой камеры CD25 с фирменным программным обеспечением.
Известно, что закладка главного источника клеток для развивающегося хрусталика глаза человека происходит из эктодермы [4]. Механизм образования хрусталикового пузырька - взаимная индукция с передней стенкой глазного бокала [3, 5]. По нашим данным, на ранних этапах развития хрусталик представлен группой клеток эктодермальной плакоды, погрузившейся за инвагинирующей передней стенкой глазного бокала. На светооптическом уровне клетки представляют собой морфологически идентичную группу с выраженной базофилией и высокими ядерно-цитоплазматическими отношениями. Хрусталиковая плакода контактирует с зоной будущей роговицы и эктодермой. Клетки в совокупности образуют плакодную пирамидку, между вершиной которой и углублением глазного бокала располагается тяж клеток, отличающихся более высокой хромофильностью по сравнению с клетками стекловидного тела. Направлены они к презумптивному месту закладки жёлтого пятна. Также наблюдается миграция переднего хрусталикового эпителия подкапсулярно, на заднем полюсе и с боковых поверхностей хрусталика внутрь пузырька мигрирует глия. Хрусталиковый эпителий хромофильный, а глия не окрашена, клетки имеют веретеновидную форму. Внутри пузырька распространяется горизонтально, а на заднем полюсе сагиттально по центру хрусталиковой подковы.
Согласно классической концепции развития хрусталика глаза человека хрусталиковая плакода после преобразования в хрусталиковый пузырёк инвагинирует дистально, и в экваториальной зоне периферические клетки, по мере погружения внутрь хрусталика, превращаются в хрусталиковые волокна [3, 15, 16]. Эта концепция не даёт представления об источниках образования задней капсулы хрусталика и особенностей её строения, сагиттальной направленности хрусталиковых волокон, не объясняет многообразие клинических форм катаракт.
По нашим данным, на 6-й неделе в хрусталике идентифицируются: 1) капсулярный эпителий переднего и экваториального полюсов; 2) выселившиеся клетки из переднего капсулярного эпителия; 3) экваториально прилежащие зоны переднего и заднего полюсов хрусталика из поперечно и сагиттально расположенной глии; 4) формирование задней глиальной мембраны; 5) начало развития сосудистой сумки хрусталика (рис. 1).
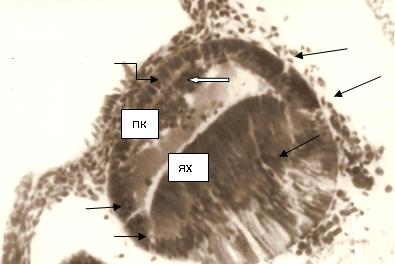
Рис. 1. Глаз эмбриона человека 6,5 недель. Микрофото. Импрегнация серебром. Ув. х400. Стрелками указаны мигранты из внутренней стенки глазного бокала. Ях - ядро хрусталика; ПК - передняя капсула. Белая стрелка указывает на выселившиеся клетки из передней капсулы хрусталика. Фигурная стрелка соответствует зоне контакта переднего капсулярного эпителия и зачатка роговицы.
На 7-й неделе все оболочки глаза и хрусталик пронизаны клетками радиальной глии. У эмбриона 7 недель идентифицируются хрусталиковая сумка и сосудистая капсула. Строма внутри пузырька состоит из глии. Мы предполагаем, что её роль заключается в координации направления хрусталиковых волокон, предотвращении ангиогенеза внутри хрусталика за счёт ингибирующих этот процесс свойств (выработка стромальными клетками гиалуроновой кислоты) [11].
Только после образования капсулы на заднем полюсе хрусталика формируется гиалоидный бассейн. Согласно литературным данным, глия в культуре вырабатывает подобно фибробластам гиалуроновую кислоту, являющуюся ингибитором ангиогенеза [9]. Это объясняет, почему сосуды капсулярной сумки не прорастают в хрусталик. Также мы нашли работы, в которых установлено, что в культуре клеток радиальная глия способна вырабатывать хрусталиковые белки кристаллины [14]. Следующее подтверждение участия глии в формировании хрусталика базируется на том, что в экспериментах с хрусталиковым пузырьком, изолированном от глазного бокала с сохранением гуморального влияния, не образуются хрусталиковые волокна. Следовательно, только контактные взаимодействия с глазным бокалом и миграция глии приводят к волокнообразованию. В отличие от Guntersson (1996) , утверждающего, что беспигментный эпителий цилиарного тела участвует в формировании заднего полюса хрусталика, в наших исследованиях закладка хрусталика и формирование заднего полюса происходит задолго до обособления фиброзной и сосудистой оболочек, а уж тем более отростков цилиарного тела. В эти сроки идентифицируется задняя глиальная капсула хрусталика, гиалоидная артерия и двухслойная эпителиальная капсула хрусталика на переднем полюсе (рис. 2а, б).
Очевидно, глия участвует в формировании всех бессосудистых структур глаза для координации в дальнейшем направления потока света; предотвращения ангиогенеза в этих структурах, способности трансформации в резидуальные АПК в условиях иммунодефицита; выработкe кристаллинов. Дополнительным подтверждением правильности нашей точки зрения является то, что не доказана выработка кристаллинов эпителием хрусталика в отсутствие глазного бокала [10].
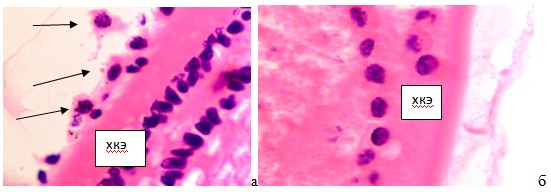
Рис. 2 а, б. Глаз плода человека 18 недель. А - передний полюс, б - зона экватора. Микрофото. Окраска г/э. Ув. х800. Идентифицируется двухслойная хрусталиковая капсула. На поверхности хрусталика располагаются макрофаги, отмеченные стрелками, способствующие обособлению хрусталика от роговицы. ХКЭ - хрусталиковый капсулярный эпителий
Таким образом, по нашим данным, идентифицировались 2 дифферона клеток в развивающемся хрусталике: эктодермальный капсулярный эпителий и глиоциты. Дополнительное подтверждение наличия второго дифферона клеток глиальной природы в формирующемся хрусталике получено с помощью реакции на белок S100, маркирующей глиоциты (bb, ll, lb).
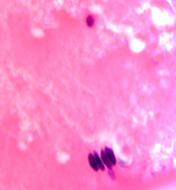
В структуре хрусталика плода нами выделены клеткиглии трёх типов:1 тип - капсулярные клетки-ядра овальной формы, крупные до 30 мкм; 2 тип - ядра круглой формы, более мелкие по сравнению с 1-м типом клеток, 15-20 мкм. 3 тип не связан с капсулой, это мигрирующий клеточный пул, находится в структуре хрусталика, идентифицированы два вида - с большим количеством цитоплазмы и узким ободком цитоплазмы (рис. 3 а, б, в).
 а
а  б
б 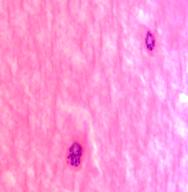 в
в
Рис. 3 а, б, в. Хрусталиковые клетки глаза плода человека 24 недель. Микрофото. Окраска г/э
Нами установлено, что взаимодействие с СТ, Р и ЦТ на поверхности хрусталика обусловлено тем, что: 1) существуют контактные участки, представленные остатками капсулярной сосудистой сумки хрусталика; 2) контактные зоны располагаются на примерно одинаковом расстоянии в 50 мкм; 3) идентифицируются якорные клетки СТ, образующие плотный контакт с капсулой хрусталика (рис. 4).
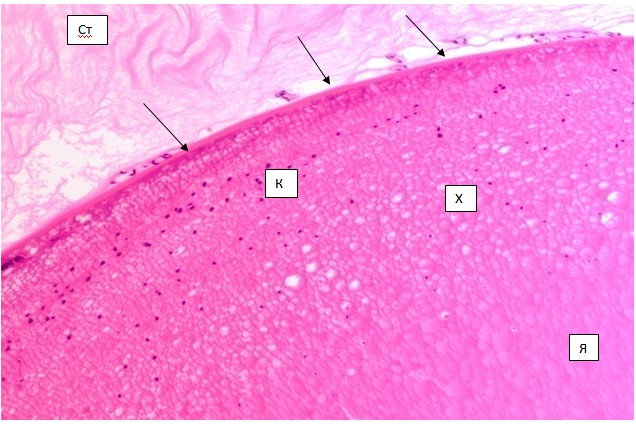
Рис. 4. Хрусталик глаза плода человека 24 недель. Микрофото. Окраска г/э. Ув. 400. Чёрными стрелками указаны остатки сосудистой капсулы хрусталика. Ст - стекловидное тело. Х - хрусталик. Я - ядро хрусталика. К - корковая зона хрусталика. Белая стрелка указывает на экваториальный обрыв капсулярного эпителия
Также нами установлена неоднородность капсулы хрусталика глаза человека. Выделены наружный гомогенный слой на всей поверхности хрусталика, внутренний с клетками на переднем полюсе и бесклеточный на заднем полюсе, и средний слой, с более бледной окраской, содержащий пустоты - следы апопотозирующих клеток второго слоя. По нашему мнению, это связано с тем, что наружный слой капсулы по всей окружности является производным глиальных клеток, выселившихся из внутренней стенки глазного бокала, и окружающий всю поверхность хрусталика (рис. 5).
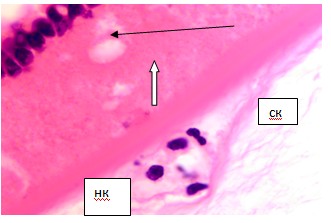
Рис. 5. Капсула хрусталика глаза плода человека 24 недель. Микрофото. Окраска г/э. Ув. х600. Чёрной стрелкой указан капсулярный эпителий; белой стрелкой - тень от апоптозирующей клетки; нк - наружный капсулярный слой; ск - сосудистая капсула
Таким образом, на основе полученных результатов сделан вывод об участии в формировании хрусталика глаза человека двух источников развития: 1) эктодермальной хрусталиковой плакоды, являющейся источником развития капсулярного эпителия; 2) глиоцитов, выселившихся из внутреннего листка глазного бокала, источник стромальных хрусталиковых клеток.
2. Сосудистая капсула хрусталика формируется после окончания миграции глиоцитов и служит поставщиком макрофагов, участвующих в обособлении хрусталика от окружающих его структур. После окончания обособления сосудистая капсула хрусталика подвергается инволюции.
3. Хрусталиковая плакода формирует хрусталиковый пузырёк, затем, в результате инвагинации, двухслойную серповидную капсулу на переднем полюсе с апоптозированием внутреннего слоя капсулы.
Рецензенты:
Красниов Ю. А., д.м.н., профессор кафедры теории и методики адаптивной физической культуры ФГАОУ ВПО «ДВФУ», г. Владивосток.
Усов В. В., д.м.н., профессор, зав. кафедрой клинической и экспериментальной хирургии ФГАОУ ВПО «ДВФУ», г. Владивосток.
Библиографическая ссылка
Рева Г. В., Гапонько О. В., Новиков А. С., Куликова К. С., Ан Е. А., Рева И. В. КОНЦЕПЦИЯ РАЗВИТИЯ ХРУСТАЛИКА ГЛАЗА ЧЕЛОВЕКА // Современные проблемы науки и образования. 2012. № 3. ;URL: https://science-education.ru/en/article/view?id=6531 (дата обращения: 15.06.2026).