


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
MONOLAYERS OF THE PURPLE MEMBRANES AS THE PLATFORM FOR BIOHYBRID NANOMATERIALS

Бактериородопсин (БР) представляет собой мембранный белок, выделяемый из пурпурных бактерий Halobacterium halobium и действующий как светозависимый протонный насос [6]. Он выгодно отличается от других светочувствительных мембранных белков своей доступностью, стабильностью и способностью ориентированно встраиваться в искусственные мембранные системы (монослои, липосомы и т.п.) [1, 6]. Мембранные системы на основе БР представляют значительный интерес как модели для исследовании молекулярных механизмов фоторецепции и как перспективные материалы для записи и хранения оптической информации, для опто- и молекулярной электроники, динамической голографии [1, 2, 7]. Метод брюстеровского рассеяния позволяет оценить толщину монослоя на границе раздела фаз, исследовать морфологию и любые структурные изменения в монослое путем измерения интенсивности рассеяния лазерного кучка, падающего на поверхность монослоя под углом Брюстера [4, 5].
Учитывая важность структурно-функциональной характеристики мембранных систем на основе бактериородопсина, целью данной работы являлось исследование фотоиндуцированных изменений в монослоях БР на границе раздела фаз жидкость/газ с помощью недавно разработанного метода брюстеровского рассеяния от поверхности монослоя.
Материалы и методы исследования
Пурпурные мембраны и БР выделяли и характеризовали по известным методикам [6]. Для сравнительной характеристики монослоя БР одновременно регистрировали изотермы поверхностного давления и брюстеровского рассеяния при равномерном сжатии монослоя подвижным барьером с постоянной скоростью 10 см2/мин. Для измерения брюстеровского рассеяния использовали следующую схему эксперимента: лазерный луч (мощность 1 мВт, длина волны 670 нм) с помощью системы зеркал направляли на монослой БР (диаметр светового пятна на монослое - порядка 1 мм) под углом Брюстера (53 °, для воздуха). Интенсивность лазерного луча, отраженного от поверхности монослоя под этим же углом, детектировали с помощью фотоумножителя (Наmamatsu R1635), а затем через систему частотных фильтров и аналоговых преобразователей записывали в компьютере для дальнейшей обработки и графического представления результатов. Вся измерительная система была смонтирована на специальной раме, расположенной на высоте 10 см над поверхностью монослоя, причем каждый из элементов оптической системы мог юстироваться независимо отдельным винтом. Такая конструкция позволяла предельно точно юстировать лазерный луч под углом Брюстера к поверхности монослоя [4, 5].
Результаты исследований и их обсуждение
Резкий подъем по изотерме поверхностное давление - площадь на молекулу БР (рис. 1, кривая 1) и высокое давление коллапса (более 45 мН/м) свидетельствуют об образовании достаточно прочных и стабильных монослоев БР на концентрированных солевых субфазах. В предыдущих исследованиях нами было показано, что наличие солей калия в водной субфазе в концентрации 100 мМ является оптимальным для формирования монослоя БР, тогда как на бидистилляте или несолевых буферах (типа трис-HCl) образование монослоя БР не происходит вовсе. Площадь на молекулу белка (А), экстраполированная из линейного участка изотермы к нулевому давлению, составляет порядка 8,0 нм2 (а при 30 мН/м - А=5,5 нм2), что соответствует данным трехмерной модели Хендерсона [3].
Изменение интенсивности брюстеровского рассеяния по мере сжатия монослоя БР (рис. 1, кривая 2) существенно отличается от такового для монослоев липидов. В монослоях БР значение интенсивности брюстеровского рассеяния увеличивается постепенно, соответственно росту поверхностного давления, тогда как для монослоев липидов характерно резкое скачкообразное увеличение брюстеровского рассеяния задолго до начала роста поверхностного давления в монослое. Достаточно резкий рост брюстеровского рассеяния наблюдается только при высоких значениях поверхностного давления (30-45 мН/м), когда домены ПМ уже организованы в конденсированный монослой.
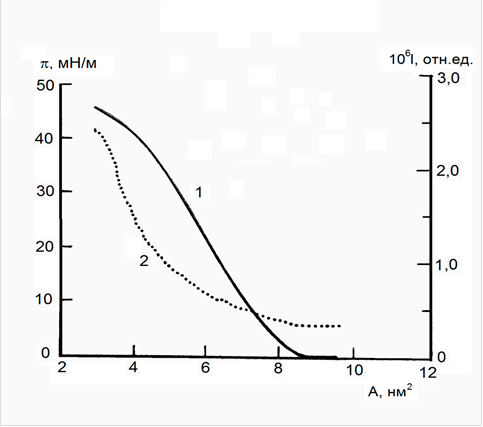
Рисунок 1. Изотермы поверхностного давления π (1) и брюстеровского рассеяния I (2), отнесенные к площади на молекулу бактериородопсина A в монослое (100 мМ KCl, 18 °C)
Максимальная интенсивность брюстеровского рассеяния монослоя БР имеет величину порядка 2,5×10-6 (отн. ед.), что существенно выше, чем для монослоев природных липидов. Используя усредненное значение показателя преломления аминокислот, равное 1,43, и соответствующие геометрические коэффициенты для установки брюстеровского рассеяния, мы рассчитали толщину монослоя БР, которая составляла 4,5 ±0,1 нм. Этот результат соответствует данным, полученным ранее на ленгмюровских пленках БР, толщина монослоя в которых составляла 4,74 ± 0,04 нм [1]. Следовательно, монослой БР на границе раздела фаз имеет толщину, характерную для нативных пурпурных мембран и эквивалентную липидному бислою. Таким образом, с помощью метода брюстеровского рассеяния было впервые непосредственно показано, что белковый монослой является адекватной моделью биологических мембран, а не «половиной» биомембраны, как в случае монослоев липидов.
Наиболее важным из полученных результатов является наблюдение обратимых изменений в брюстеровском рассеянии монослоев бактериородопсина при чередовании облучения светом с длиной волны 570 нм и темновой релаксации (или облучения светом с длиной волны 420 нм). Свет, предназначенный для активации молекул БР, с помощью гибкого кварцевого волновода фокусировали вертикально на монослой ПМ точно в месте падения (под углом Брюстера) зондирующего зерного луча. В такой геометрии свет, предназначенный для активации молекул БР, не попадал в фотоумножитель, регистрировавший только интенсивность брюстеровского рассеяния лазерного луча. Вся установка была заключена в затемненный кожух для предотвращения попадания постороннего излучения на монослой БР.
Абсолютные значения обратимых изменений в брюстеровском рассеянии монослоев бактериородопсина (рис. 2) хорошо воспроизводятся в серии последовательных измерений (с интервалом возбуждение - релаксация, равным 1 мин); существенно зависят от интенсивности света; расстояния между светопроводящим оптическим волокном и поверхностью монослоя, а также от величины поверхностного давления во время эксперимента. Так, например, изменения находятся практически в области шумов для растянутых монослоев ПМ (поверхностное давление ниже 1,0 мН/м), резко увеличиваются с ростом поверхностного давления, достигая максимальных значений при 30 мН/м, и уменьшаются при более высоких давлениях в области коллапса монослоя.
Эффекты, изображенные на рис. 2, очень похожи на те, которые были обнаружены при исследовании изменений оптических и фотоэлектрических свойств БР в ленгмюровских пленках при фотоактивации [1]. Принимая во внимание, что интенсивность брюстеровского рассеяния уменьшается при облучении светом с длиной волны 570 нм и восстанавливается при темновой релаксации (рис. 2), можно сделать вывод, что толщина монослоя ПМ уменьшается или увеличивается соответственно. Эти изменения можно объяснить как реорганизацией самих белковых молекул в тримере [1, 2, 6, 7], так и их спиралей при фотоактивации в более «растянутое» состояние.
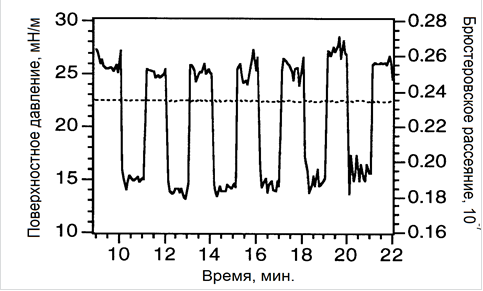
Рисунок 2. Интенсивность брюстеровского рассеяния I (1) и поверхностное давление π (2) монослоев бактериородопсина на водной субфазе 100 мМ KCl при включении и выключении облучения (570 нм).
Заключение
Таким образом, впервые было показано, что толщина монослоя ПМ непосредственно на границе раздела жидкость/воздух хорошо соответствует толщине нативных биомембран. Это позволяет считать монослои ПМ адекватной моделью пурпурных мембран. Авторы надеются, что полученные данные по обратимым фотоиндуцированным изменениям размеров монослоя ПМ будут способствовать более глубокому пониманию процессов фоторецепции в биомембранах и могут быть использованы при создании биосенсорных устройств.
Рецензенты:
- Царькова Марина Сергеевна, доктор химических наук, профессор кафедры органической и биологической химии ФГБОУ ВПО «Московская государственная академия ветеринарной медицины и биотехнологии имени К. И. Скрябина» (ФГБОУ ВПО МГАВМиБ), г. Москва.
- Максимов Владимир Ильич, доктор биологических наук, профессор, профессор кафедры физиологии животных ФГБОУ ВПО «Московская государственная академия ветеринарной медицины и биотехнологии им. К. И. Скрябина» (ФГБОУ ВПО МГАВМиБ), г. Москва.
Библиографическая ссылка
Зайцев С.Ю., Соловьева Д.О. МОНОСЛОИ ПУРПУРНЫХ МЕМБРАН КАК ОСНОВА БИОГИБРИДНЫХ НАНОМАТЕРИАЛОВ // Современные проблемы науки и образования. 2012. № 3. ;URL: https://science-education.ru/en/article/view?id=6386 (дата обращения: 22.06.2026).