


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
HOMOLOGY AT SPECIES OF WHEAT IN CONNECTION WITH PHENOMOGENOMICS OF THEIR QUANTATIVE SIGNS

Цель настоящих исследований - с помощью феномогеномных исследований продукционных признаков видов-гомологов убедиться в различных уровнях «феномности» этих признаков у T.miguschovae AbGD и T.aestivum AuBD. Далее, основываясь на результатах феномогеномных исследований, попытаться получить полный гомолог мягкой пшеницы.
Методика. Для этого, с использованием коэффициента удельной продуктивности (далее Ку.п.), был подтвержден двухфеномный уровень продуктивности видов гомологов T.kiharae и T.spelta, а также у T.miguschovae и трёхфеномный у T.aestivum. На основании полученных результатов признано, что T.miguschovae неполный гомолог мягкой пшеницы. Один из возможных способов создания полного гомолога - это попытаться реализовать вклад третьего элементарного диплоидного генома у гексаплоидных Т.miguschovae или Т.kiharae при помощи мутагенеза. В результате обработки семян этих видообразцов супермутагеном N-нитрозо-N-метилмочевиной (далее НММ) из Т. kiharae получен полный гомолог, что подтверждено иммунохимическим и электрофоретическим анализами, проведенными в лаборатории биохимии ВИР (Санкт-Петербург).
Результаты и обсуждение. Ку.п. отражает соотношение зерна воспроизведенным продуктивным побегом к массе этого же побега без зерна, поскольку наименьшей единицей, в которой реализуется взаимосвязь вегетативных и генеративных сфер растения, является продуктивный побег [6]. Данный показатель, который очень близок к коэффициенту хозяйственной эффективности урожая (далее Кхоз), вполне объективно отражает продуктивность пшеницы.
Из данных табл. 1 видно, что Ку.п. T.kihаrae и T.spelta практически одинаков и значительно ниже, чем у представителей мягкой пшеницы. Ку.п. гомолога мягкой пшеницы T.miguschovae (0,51) также на уровне гексаплоидных T.kiharae и T.spelta.
Таблица 1 - Удельная продуктивность видов-гомологов (1989 г.)
|
Видообразец |
№ по кат. ВИР, сорт |
Геном |
Масса, г |
К у.п. |
|
|
зерна |
вегетативной части побега |
||||
|
T.spelta |
19097 |
AuBD |
1,48 |
2,81 |
0,53 |
|
53347 |
AuBD |
0,85 |
1,69 |
0,50 |
|
|
T.kiharae |
47897 |
AbGD |
0,67 |
1,41 |
0,48 |
|
T.aestivum |
Найя |
AuBD |
1,44 |
1,84 |
0,78 |
|
Бирюсинка |
AuBD |
0,87 |
1,16 |
0,75 |
|
|
Донщина |
AuBD |
1,74 |
2,18 |
0,80 |
|
|
T.miguschovae |
57361 |
AbGD |
0,71 |
1,40 |
0,51 |
|
НСР05 |
|
|
|
|
0,07 |
Учитывая, что в среднем Ку.п. T.aestivum 0,78, вычисляем вклад в данный признак одного диплоидного генома или феном. Для чего 0,78 : 3 = 0,26. Уровень феномности T.kiharae и T.spelta можно показать, разделив их признаки на выявленный вклад диплоидного генома: 0,48 : 0,26 = 1,85≈2,0 и 0,52 : 0,26 = 2,0. Значит, T.kiharae и T.spelta обладают двухфеномным уровнем данного признака, и у них не проявляется вклад одного из трёх диплоидных геномов. Однако и гомолог мягкой пшеницы T.miguschovae также обладает двухфеномным уровнем: 0,51 : 0,26 = 1,96 ≈ 2,0.
При продолжении исследования Ку.п. получены аналогичные результаты (табл. 2). По крайней мере, из данных табл. 2 однозначно следует, что спельтоидные виды - гомологи T.kiharae и T.spelta, несмотря на свой гексаплоидный статус, имеют двухфеномный уровень продуктивности.
Таблица 2 - Удельная продуктивность видов гомологов (1990 г.)
|
Видообразец |
№ по кат. ВИР, сорт |
Геном |
Масса, г |
Ку.п. |
|
|
зерна |
вегетативной части побега |
||||
|
T.spelta |
19097 |
AuBD |
1,25 |
2,52 |
0,50 |
|
T.kiharae |
47897 |
AbGD |
0,93 |
1,93 |
0,47 |
|
// -- // -- // |
// -- // -- |
0,61 |
1,30 |
0,47* |
|
|
T.aestivum |
Донщина |
AuBD |
1,11 |
1,36 |
0,82 |
|
Бирюсинка |
// -- // |
0,94 |
1,28 |
0,73 |
|
|
// -- // -- // |
// -- // |
0,59 |
0,79 |
0,75* |
|
|
Найя |
// -- // |
1,65 |
2,13 |
0,77 |
|
|
Фури |
// -- // |
1,46 |
1,90 |
0,77 |
|
|
Моск. 35 |
// -- // |
1,99 |
2,84 |
0,70 |
|
|
Свенно |
// -- // |
1,73 |
2,41 |
0,72 |
|
|
Ролло |
// -- // |
1,72 |
2,36 |
0,73 |
|
|
T.miguschovae |
57361 |
AbGD |
0,69 |
1,39 |
0,50 |
|
НСР05 |
|
|
|
|
0,08 |
* - те же видообразцы, выращенные в других условиях.
Вместе с тем гомолог мягкой пшеницы T.miguschovae по своим продукционным показателям соответствует T.spelta и T.kiharae. У него Ку.п. 0,50, а у представителей мягкой пшеницы в среднем 0,75. Следовательно, T.miguschovae неполный гомолог мягкой пшеницы, что следует из соотношения, согласно закону гомологических рядов:
Т. spelta AuBD(2) ≈ (2)AbGD T. kiharae,
Т. aestivum AuBD(3) ≠ (2) AbGD T. miguschovae
(в скобках продуктивность, выраженная в феномах). На неполную гомологию Т.miguschovae с T.aestivum указывает двухфеномный уровень его продуктивности, что до настоящего времени не принималось в расчет. Полная же гомология определяется одинаковым или близким фенотипическим эффектом [7].
Для достижения этого одинакового или близкого фенотипического эффекта (полной гомологии с мягкой пшеницей) необходима реализация вклада третьего диплоидного генома или у Т. miguschovae, или Т. kiharae. Исходя из этих соображений, семена данных образцов обработаны НММ. Обработанные семена высеяли на подготовленной грядке вдали от посевов. Уже в М1 среди образцов Т.miguschovae появились растения со светлыми колосьями и типичные Т.kiharae. У исходных форм Т.kiharae существенных морфологических изменений не наблюдалось.
В М2 у Т.kiharae, наряду с формами, похожими на неё, появились растения с темноокрашенным и более плотным колосом, то есть напоминающие Т.miguschovae, а также очень близкие к Т.spelta. Среди мутантных растений изначального Т.miguschovae в М2 было обнаружено растение, похожее на Т.kiharae, но отличавшееся более крупными размерами листьев и меньшей их опушенностью. Колосья его имели сизоватый восковой налет, как и у сортообразцов мягких пшениц. Семена с этих растений были высеяны отдельно в теплице. Остальные семена М2 Т.kiharae и T.miguschovae высеяли поздней осенью, опять же на специально подготовленном участке, вдали от производственных посевов. Высевая под зиму, мы пытались усилить мутагенный эффект [5]. В М3 из семян мутантного растения с крупными и малоопушенными листьями взошли только два. При созревании они оказались типичными Т.timopheevii.
Однако в М3 точно в одном из рядков перезимовавшего Т. kiharae было обнаружено растение, по внешнему виду соответствующее Т.аestivum. При пересевах выяснилось, что в отличие от исходного T.kiharae они имели озимый образ жизни и, как большинство представителей второй филогенетической ветви, выделялись более интенсивной темно- зеленой окраской.
Таким образом, нами был получен предполагаемый полный гомолог мягкой пшеницы [8]. В вегетационном опыте было выявлено, что по зерновой продуктивности он был на уровне сортообразцов мягкой пшеницы и на 1/3 превосходил Т.kiharae, тo есть обладал «трехфеномной» продуктивностью (табл. 3).
Таблица 3 - Продуктивность полного гомолога по сравнению с исходными видами, г/сос
|
Вид |
2n |
Уровни минерального питания |
В ср. |
||||
|
К |
N0.05 |
N0.10 |
N0.20 |
N0.60 |
|||
|
T.timopheevii |
28 |
2,1 |
11,5 |
10,6 |
8,7 |
4,9 |
7,6 |
|
T.kiharae |
42 |
2,0 |
10,2 |
12,8 |
11,6 |
6,0 |
8,5 |
|
Полный гомолог |
42 |
3,6 |
12,6 |
17,5 |
19,8 |
9,6 |
12,6 |
|
НСР05 |
|
|
|
|
|
|
1,2 |
К - контроль без удобрений; N г. д.в./сос.
Из данных табл. 3 видно, что различия по продуктивности между T.timopheevii и T.kiharae не существенны, тогда как между последним и полным гомологом различия на всех вариантах достоверно отличаются в пользу полного гомолога.
Что характерно, и в среднем продуктивность T.kiharae на уровне T.timopheevii. Разница между полным гомологом и T.kiharae (12,6г - 8,5г = 4,1г) составляет 32% или 1/3. Учитывая двухфеномную продуктивность T.kiharae, вычисляем вклад диплоидного генома в его признак: 8,5 : 2 = 4,25 получаем вклад диплоидного генома, который соответствует разнице между T.kiharae и полным гомологом. Далее определяемся с уровнем феномности признака полного гомолога, который равняется 12,6 : 4,25 = 2,96≈3,0.
Иммунохимический анализ, проведенный в лаборатории биохимии ВНИИР им. Н.И. Вавилова (г. Санкт-Петербург), несмотря на чрезвычайное сходство полного гомолога с T.aestivum, показал наличие у него генома Ab, тогда как у истинной мягкой пшеницы Au (фото 1).

Фото 1. Иммунохимический анализ (двойная иммунодиффузия) липопротеинов:
1 - T.boeoticum; 2 - полный гомолог; 3 - T.timopheevii, выщепившийся из T.miguschovae; 4 - T.timopheevii; 5 - T.militinae; 6 - T.urartu; 8 - T.aestivum; 7, 9, 10 - мутанты полученные из T.militinae.
На фото 1 четко видна одинаковая реакция T.urartu Au и T.aestivum AuBD, в отличие от представителей второй филогенетической ветви, включая и полный гомолог, у которых геном Ab. В то же время мутантные формы тетраплоидного T.militinae и T.timopheevii, выщепившийся из Т. miguschovae, четко реагировали на Ab. Аналогично реагирует полный гомолог, несмотря на то что по фенотипу он - копия мягкой пшеницы. Однако у полного гомолога вместо генома G обнаруживались признаки генома В. Присутствие генома D у полного гомолога, так же как и у образцов мягкой пшеницы, использованных в анализах, подтверждается.
Был проведен электрофоретический анализ глиадинов этих же видов пшениц с использованием методики [1]. Из представленной фореграммы видно, что спектры глиадинов филогенетически близких видов T.militinae и T.timopheevii имеют практически идентичный состав компонентов, который сильно отличается от спектра мягкой пшеницы (фото 2).
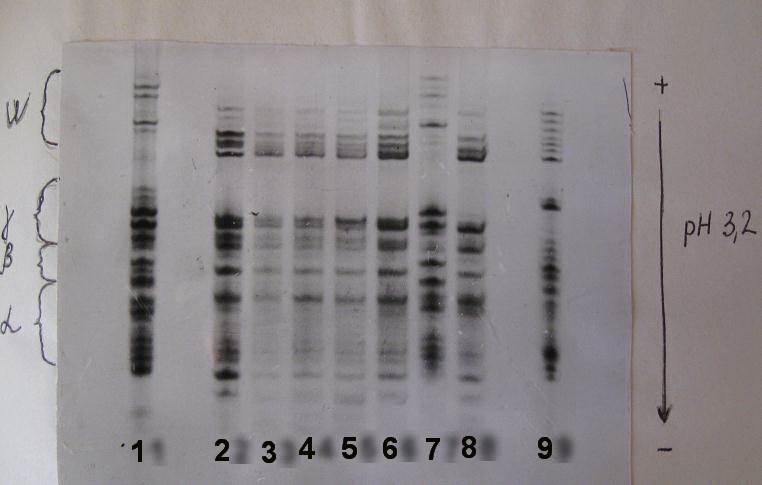
Фото 2. Электрофорез глиадинов исследуемых образцов:
1 - T.aestivum; 2 - T.militinae; 3, 4, 5 - мутанты из T.militinae; 6 - T.timopheevii, выщепившийся из T.miguschovae; 7 - полный гомолог; 8 - T.timopheevii; 9 - T.durum.
Полный гомолог выделяется оригинальным промежуточным типом спектра, в котором присутствуют как компоненты, характерные для пшениц ряда T.timopheevii, так и в большей степени компоненты, специфичные для мягкой пшеницы. Очевидно, трансформация генома G в геном В предопределила фенотипическое сходство полного гомолога с мягкой пшеницей T.aestivum. О возможности таких трансформационных изменений говорит тот факт, что геном G может являться производным генома В [3].
Поскольку полный гомолог фенотипически идентичен мягкой пшенице, то для удобства его можно обозначить, как T.aestivum TM (где ТМ указывает на его происхождение из пшениц ряда Тимофеева). Более того, феноменальное сходство с мягкой пшеницей позволило дать ему и сортовое название Рассвет-1. Отсюда гомологию изучаемых гексаплоидные видов можно представить следующим образом:
T.spelta AuBD (2) ≈ (2) AbGD T.kiharae,
T.aestivum L. AuBD (3) ≈ (3) AbBD T.aestivum TM.
Учитывая, что из T.spelta в результате радиационного воздействия получали мягкую пшеницу и, наоборот, из мягкой пшеницы выщеплялась спельта, получение полного гомолога представляется вполне закономерным [6]. Очевидно, что если из T.spelta появляется T.aestivum, то выщепление из гомологичного спельтоидного T.kiharae, полного гомолога с трёхфеномным уровнем продукционных признаков, фенотипически повторяющего мягкую пшеницу, достаточно логично и согласуется с законом гомологических рядов. В принципе, без учёта дифилетической природы происхождения изучаемых пшениц, можно считать, что был произведен ресинтез мягкой пшеницы.
Вероятно, благодаря морфофизиологическому и генотипическому сходству с мягкой пшеницей полный гомолог Рассвет-1 уже не проявлял такую же устойчивость к бурой ржавчине, как исходный вид. Хотя относительно сортообразцов мягкой пшеницы он выглядел предпочтительнее.
То есть близость или сходство полного гомолога Рассвет-1 с мягкой пшеницей, как в генетическом, так и фенотипическом плане, негативно сказывается на его устойчивости к бурой ржавчине. Вместе с тем некоторые генетические отличия Рассвет-1 делают его ценным исходным материалом для селекционной практики. Так при гибридизации Рассвет-1 с сортом мягкой пшеницы Юбилейная 100, с коротким вегетационным периодом, получена довольно скороспелая и относительно высокопродуктивная форма.
Заключение. Благодаря феномогеномным исследованиям видов-гомологов установлено, что T.miguschovae из-за своего двухфеномного уровня количественных признаков не является полным гомологом мягкой пшеницы T.aestivum, у которого, как и должно, трёхфеномный уровень аналогичных признаков, включая и зерновую продуктивность. Полный гомолог Рассвет-1, практически идентичный мягкой пшенице, получен из T.kiharae, после соответствующего воздействия на его семена НММ. Он является ценным исходным селекционным материалом для практической селекции.
Рецензенты:
Усатов А.В., д.б.н., профессор, заведующий отделом изменчивости ген, НИИ биологии Южного федерального университета. Министерство образования РФ, ГБОУ ВПО «Южный федеральный университет», г. Ростов-на-Дону.
Титаренко А.В., д.с.-х.н., старший научный сотрудник, заведующий отделом селекции и семеноводства зерновых и зернобобовых культур Донского НИИ сельского хозяйства, Министерство сельского хозяйства РФ, ГНУ Донской НИИСХ Россельхозакадемии, Ростовская область, Аксайский район, п. Рассвет.
Библиографическая ссылка
Романов Б.В., Авдеенко А.П. ГОМОЛОГИЯ У ВИДОВ ПШЕННИЦЫ В СВЯЗИ С ФЕНОМОГЕНОМИКОЙ ИХ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ // Современные проблемы науки и образования. 2012. № 1. ;URL: https://science-education.ru/en/article/view?id=5586 (дата обращения: 29.07.2026).