


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
SOMACLONAL VARIABILITY IN THE ADAPTATION OF WOLFFIA ARRHIZA (L. ) HORKEL EX WIMMER PLANTS TO THE EFFECTS OF SYNTHETIC HERBICIDE 2,4-D

Цель данной работы - исследование сомаклональной изменчивости вольфии бескорневой Wolffia arrhiza (l.) Horkel ex Wimmer, относящейся к семейству рясковых. Представители семейства рясковых широко распространены в водоемах с невысокой скоростью течения, включая непроточные. Они характеризуются относительной простотой строения, высокой скоростью размножения, преимущественно вегетативного, которое сопровождается быстрым накоплением биомассы. Известно, что представители семейства отличаются устойчивостью к целому ряду ксенобиотиков. Во многих работах показано, что различные виды рясковых могут расти в водных системах с высокой степенью антропогенного загрязнения и участвовать в процессах биодеградации органических поллютантов различной природы и/или аккумуляции ионов тяжелых металлов с высоким индексом обогащения [4, 5, 7]. С другой стороны, поглощение и накопление некоторых токсических веществ приводит к физиологическим и выраженным фенотипическим изменениям этих растений, не вызывая при этом их гибели, на чем основано использование различных видов рясок в качестве объектов для проведения биотестов [6, 10]. При этом показано, что внутри одной популяции растения могут в значительной степени различаться по степени устойчивости к действию внешних факторов [8, 9]. Подобная вариабельность в пределах одного биотопа определяется генетической и фенотипической гетерогенностью природных популяций и лежит в основе устойчивости популяции к изменению условий обитания [2].
Материалы и методы
В работе исследовали сомаклональную фенотипическую изменчивость вольфии бескорневой под воздействием синтетического гербицида ауксинового ряда - 2,4-D, обладающего мутагенными свойствами. Поверхностно стерилизованные растения выращивали на среде Гамборга - Эвелега В-5 с разной степенью разведения: в 5 раз (1/5 среды В-5) и в 50 раз (1/50среды В-5) при 24 оС с искусственным освещением интенсивностью 6000 люкс при 16-ти часовом фотопериоде.
Для получения растений с измененным фенотипом в состав среды асептически вводили раствор 2,4- D в диапазоне концентраций от 1мкМ до 100мкМ и культивировали растения в течение 3 недель, как при 16-ти часовом фотопериоде, так и в темноте, после чего осуществляли первичный анализ полученных форм. Растения с измененным фенотипом пересаживали на свежую питательную среду и культивировали в оптимальном для них температурном режиме и фотопериоде. Для культивирования обесцвеченных клонов использовали жидкие и агаризованные минеральные среды, содержащие 0,5-1 % сахарозы.
Эксперименты проводили в 3-х биологических и 3-х аналитических повторностях. Статистическую обработку проводили с помощью программы Microsoft Excel 2003. На рисунке представлены средние арифметические значения результатов, полученных в отдельных экспериментах и их стандартное отклонение от среднего значения.
Полученные результаты и их обсуждение
Культивирование растений вольфии в присутствии 2,4-D показало, что введение в среду 2,4-D в концентрации 1 - 20 мкМ приводит к заметному ускорению вегетативного размножения и к значительному приросту сырой биомассы. Однако при увеличении концентрации до 50 - 100 мкМ наблюдается ингибирование роста и развития растений, что проявляется в торможении процесса вегетативного размножения - почкования, вплоть до его полного прекращения, а также появлении мутантных форм растений. При этом высокие дозы ауксина приводили к гибели основной части растений, а среди выживших растений формировались фенотипически измененные формы.
При анализе вариабельности культуры было показано, что в среднем до 20 % растений в популяции проявляют фенотипическую изменчивость. Среди наблюдаемых фенотипических вариаций можно выделить три основных типа, которые представлены на рисунке 1.
Первый тип характеризуется изменением средних размеров растений вольфии. Были получены макро- и микроформы, размеры которых были соответственно в 2 раза больше и в 1,5 раза меньше размеров исходных растений. Стабильность макро- и микроформ в процессе длительного культивирования существенно отличалась, что позволяет предполагать участие разных механизмов в их образовании. Микроформы реверсировали в процессе нескольких пассажей, тогда как увеличенные размеры растений сохранялись в течение многих поколений без изменения способности растений к миксотрофному (фото-гетеротрофному) типу питания. Кроме того, эти растения отличались и тканевой морфологией: у них четко выражены "верхняя пластинка" - зона, где сконцентрированы фотосинтезирующие клетки, а также слабоокрашенная, сильно вакуолизированная вентральная зона.
Второй тип сомаклонов характеризуется образованием конгломератов (агрегатов) неразделившихся растений. Данный тип сохранялся при многократных пассажах и зачастую сопровождался изменением формы отдельных растений в агрегатах - появлялись удлиненные растения, а также наблюдались "размытые", нечеткие границы отдельных растений, входящих в конгломерат. Вероятно, в этом случае могли быть затронуты механизмы биосинтеза полимеров клеточных стенок растений, либо регуляции экспрессии гликолитических ферментов, определяющих собственно разделение «материнских» и «дочерних» растений.


а) б)


в) г)
Рис. 1. Сомаклональная изменчивость W. arrhiza:
а) тип 1 - линия с измененными размерами (слева - макроформа, справа - исходный фенотип);
б) тип 2 - линия с измененным синтезом клеточных стенок (конгломераты растений);
в, г) тип 3 - линии растений, дефектные по синтезу пигментов
Третий тип изменений затрагивает механизм биосинтеза пигментного аппарата и проявляется в уменьшении содержания хлорофилла и формировании желтых и белых форм растений. При изменении условий культивирования - пересадке на свежую среду - у части таких клонов растений наблюдалась частичная спонтанная реверсия до слабоокрашенных форм, но она была временной и исчезала в процессе роста культуры. Однако эти изменения не затрагивают способности вольфии к вегетативному размножению и гетеротрофному питанию.
Получение искусственных сомаклонально измененных форм растений позволило выявить несколько закономерностей.
Во-первых, показано, что чувствительность вольфии к действию 2,4-D зависит от освещенности. Так, при культивировании на свету была отмечена большая токсичность ауксина: концентрация 20 мкМ обеспечивала ингибирование вегетативного размножения растений по сравнению с концентрациями 1-10 мкМ, а последующее увеличение содержания 2,4-D в среде культивирования приводит к тому, что выживают единичные растения. В темноте токсичность 2,4-D была гораздо менее выражена и не приводит к массовой гибели растений. Образование неразделившихся форм происходит преимущественно на свету (дозовый порог ауксина при этом ниже, а процент выхода - больше), тогда как образование крупных растений - выше при культивировании в темноте. Возможно, это связано с различиями в действии препарата при различных типах метаболизма.
Вторая закономерность заключается в том, что измененные формы проявляются преимущественно при перенесении растений на свежую (минеральную) среду культивирования, мы условно обозначили как «отдаленный эффект». Тот факт, что фенотипические изменения наблюдаются у целых растений и сохраняются на протяжении многих поколений и пассажей, предполагает, что изменениям подвергаются отдельные инициальные меристематические клетки, формирующие измененные растения после деления почкованием.
Далее исследовали зависимость развития полученной линии макроформ вольфии от обеспеченности элементами минерального питания, которая отражает особенности взаимодействия данной популяции со средой обитания в сравнении с исходным фенотипом. Для этого проводили длительное культивирование на среде Гамборга - Эвелега В-5 с разной степенью разведения: в 5 раз (1/5 среды В-5) и в 50 раз (1/50среды В-5).
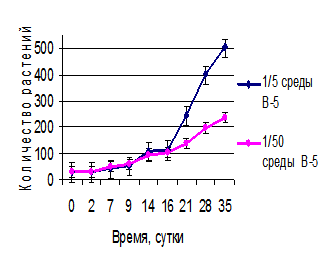
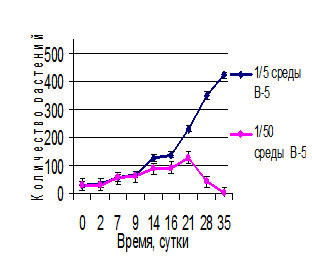
Полученные экспериментальные данные приведены на рисунке 2. На ранних этапах развития общая численность обеих популяций увеличивается на обеих средах, причем по скорости вегетативного размножения макроформа превосходит исходный фенотип. Вероятно, при росте на оптимальной минеральной среде происходит быстрое накопление биополимеров, необходимых для развития. При этом сокращается время, необходимое для генерации новых вегетативных почек и формирования молодых растений. Однако по мере истощения питательной среды скорость деления макроформы начинает замедляться и после трех недель культивирования уступает родительской форме. Надо заметить, что длительность периода генерации новых почек может зависеть и от внутрипопуляционных взаимодействий между растениями, которые опосредуются экскрецией биологически активных метаболитов. Однако эти аспекты требуют дополнительных исследований.
а) б)
Рис. 2. Развитие популяции растений W. arrhiza при разной обеспеченности элементами минерального питания: а) исходный фенотип; б) макроформа
Еще более ярко этот феномен проявляется на обедненной питательной среде. В условиях минимального минерального питания (на 1/50 среды В-5) дольше развивается исходная форма. При этом все растения мутантной формы погибают к 35-ому дню культивирования. Таким образом, можно сделать вывод, что исходная культура лучше приспособлена к среде с малым количеством питательных веществ по сравнению с макроформой, тогда как растения макроформы характеризуются повышенной потребностью в минеральном питании.
Полученные данные позволяют рассматривать макрокультуру вольфии бескорневой как перспективный объект для разработки технологий фиторемедиации водоемов. Высокая скорость размножения при быстром накоплении биомассы на богатых минеральными и органическими веществами эвтрофных водоемах будет способствовать быстрой утилизации нетоксичных загрязнений. При этом по мере очистки («истощение среды») скорость размножения популяции макроформы вольфии заметно снижается, количество растений уменьшается, что может облегчать их сбор и удаление.
Заключение
Способность к образованию устойчивых жизнеспособных форм под воздействием внешних факторов указывает на высокую морфогенетическую пластичность популяции растений вольфии бескорневой, в основе которой лежит генетическая гетерогенность. Образование нескольких фенотипически измененных линий растений вольфии можно рассматривать как проявление универсального механизма адаптации популяции водных растений к действию ксенобиотиков, приводящего к более полному проявлению адаптивной стратегии [2]. Например, образование макроформ с повышенными трофическими потребностями и высокой продуктивностью может обеспечивать более быструю и эффективную детоксикацию поллютантов, в том числе за счет их использования в качестве источника углерода при переходе на гетеротрофное питание.
Таким образом, полученные данные позволяют охарактеризовать вольфию бескорневую как весьма перспективный биотехнологический источник получения селекционных форм для фитобиоценозов с высокой продуктивностью или использования таких форм для биоремедиации водоемов и в целях биотехнологического производства.
Рецензенты:
Бородулин В.Б., профессор, д.м.н., зав. кафедрой биохимии ГБОУ ВПО „Саратовский государственный медицинский университет имени В. И. Разумовского" Министерства здравоохранения и социального развития Российской Федерации, г. Саратов.
Карпунина Л.В., профессор, д.б.н., профессор кафедры микробиологии, вирусологии и иммунологии ФГБОУ ВПО «Саратовский государственный аграрный университет имени Н.И.Вавилова» (ФГОУ ВПО «Саратовский ГАУ») (Министерство сельского хозяйства РФ), г. Саратов.
Библиографическая ссылка
Галицкая А.А., Соколов О.И., Селиванова О.Г., Селиванов Н.Ю. СОМАКЛОНАЛЬНАЯ ИЗМЕНЧИВОСТЬ ПРИ АДАПТАЦИИ РАСТЕНИЙ WOLFFIA ARRHIZA (L. ) К ВОЗДЕЙСТВИЮ СИНТЕТИЧЕСКОГО ГЕРБИЦИДА 2,4–D // Современные проблемы науки и образования. 2012. № 1. ;URL: https://science-education.ru/en/article/view?id=5531 (дата обращения: 29.07.2026).