


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
RESULTS OF A STUDY OF THE NUMBER AND CONDITION OF FATTY LIVER CELLS IN RATS WITH HYPOTHERMIA

Тучные клетки (ТК) являются одними из наиболее хорошо изученных клеток иммунной системы. Они происходят от плюрипотентных CD34+/CD117+ стволовых клеток костного мозга [1]. После миграции из костного мозга незрелые ТК циркулируют по микроциркуляторному руслу, пока не достигают периферических тканей, где созревают и подвергаются росту и дифференцировке [2; 3].
ТК в небольшом количестве постоянно присутствуют в печени и располагаются преимущественно в портальных трактах возле сосудов и желчных протоков. Эти клетки имеют очень важное значение при повреждении печеночной ткани. После воздействия повреждающих факторов холангиоциты начинают активно синтезировать фактор стволовых клеток (SCF), который является мощным хемоаттрактантом и вызывает миграцию ТК в печень. Интересно отметить, что после гепатэктомии 70% ткани печени с течением времени уровень SCF увеличивается и достигает максимума через 7 дней [4]. Мигрировавшие в печень ТК активируются, их количество возрастает, они дегранулируют, секретируя множество медиаторов, таких как гепарин, гистамин, триптаза, химаза, TNF, TGF-β1, TNF-α, IL-33, цитокины, основной фактор роста фибробластов (bFGF), лейкотриены и матриксные металлопротеиназы, которые активируют регенерацию, ангиогенез и пролиферацию гепатоцитов [5; 6].
Рутинное окрашивание гематоксилином и эозином дает мало информации об активности и морфологии ТК, а окраска толуидиновым синим лишь позволяет оценить количественные характеристики ТК и степень их дегрануляции. Поэтому была разработана комбинированная окраска ТК альциановым синим и сафранином О. Достоинством данного метода является то, что с помощью него можно дифференцировать молодые и зрелые ТК на основе содержания гепарина. При этом молодые незрелые ТК, содержащие слабо сульфатированные гликозаминогликаны (предшественники гепарина), окрашиваются в синий цвет, а зрелые и активные, за счет высокого содержания гепарина, окрашиваются в красный цвет [7]. Поэтому с помощью данного метода можно выявить незрелые ТК, мигрировавшие из микроциркуляторного русла.
В настоящее время имеются доказательства того, что ТК являются важными клеточными регуляторами при заболеваниях печени человека. Их число увеличивается, и они ремоделируют печеночную ткань при первичном билиарном холангите, первичном склерозирующем холангите, обструкции желчных протоков, гепатите, повреждении печени, вызванном алкоголем, стеатозе, стеатогепатите, фиброзе и циррозе печени, печеночноклеточном раке и холангиокарциноме, отторжении печени при трансплантации печени [8].
Гепарин, высвобождающийся из ТК при их активации, способен вызывать важные гомеостатические эффекты, ограничивающие развитие воспаления и способствующие регенерации поврежденной ткани [9]. Показано, что этот мукополисахарид, связываясь со многими белками, вовлеченными в различные стадии воспалительного каскада, проявляет множество эффектов, в том числе: стимулирует фибринолитическую и антикоагулянтную активность, ингибирует рекрутирование провоспалительных клеток в очаг повреждения и адгезию нейтрофилов к эндотелиальным клеткам сосудов, а также стимулирует ангиогенез [10; 11].
Участие ТК в регенерации органов после воздействия различных стрессорных повреждающих факторов, и в том числе гипотермии, активно изучается [12-14]. В то же время функциональная активность ТК в адаптации и регенерации печени после воздействия холодового стресса изучена недостаточно [15].
Целью работы было исследование количества и состояния тучных клеток в печени экспериментальных животных после воздействия однократной глубокой иммерсионной гипотермии.
Материалы и методы исследования. Исследование выполнено на кафедре судебной медицины имени профессора В.Н. Крюкова и патологической анатомии с курсом ДПО ФГБОУ ВО «АГМУ» Минздрава России. Исследование одобрено локальным этическим комитетом ФГБОУ ВО «АГМУ» Минздрава России (протокол № 3 от 30.10.2024 г.). Эксперимент выполнен на 30 6-месячных здоровых крысах-самцах Wistar с массой тела 255±15 граммов, выращенных в виварии Федерального государственного бюджетного научного учреждения «Федеральный исследовательский центр Институт цитологии и генетики Сибирского отделения Российской академии наук» (г. Новосибирск, Российская Федерация). Глубокую иммерсионную гипотермию (ГИГ) моделировали, помещая животных в индивидуальных клетках в воду температурой 5 °С, при температуре окружающего воздуха 7 °С. Критерием достижения глубокой степени иммерсионной гипотермии являлась ректальная температура 20-25 °С. Время экспозиции в среднем составляло 40±7 мин. Животные были подвергнуты рандомизации с выделением 5 экспериментальных групп по 5 крыс: 1-я группа – животные выводились из эксперимента декапитацией сразу после прекращения охлаждения; 2-я группа – через 2 суток; 3-я группа – через 7 суток; 4-я группа – через 14 суток и 5-я группа – через 30 суток. Животных умерщвляли цервикальной дислокацией. В качестве наркоза перед цервикальной дислокацией применяли хлороформ. Группой контроля служили 5 интактных животных.
Для гистологического исследования образцы ткани печени забирались из правой боковой, левой боковой, правой центральной и левой центральной долей. Фиксировали фрагменты печени в 10% нейтральном формалине и жидкости Карнуа в течение 24 часов, затем в автомате TISSUE-TEK VIPTM6 (Sakura, Япония) осуществляли проводку материала по изопропиловому спирту и просветляли в ксилоле. Заливку в парафин осуществляли в станции парафиновой заливки TISSUE-TEK TEC 5 (Sakura, Япония). Для заливки использовали парафин Histomix (BioVitrum, Россия). Серийные срезы толщиной 5–7 мкм изготавливали на роторном микротоме Accu-Cut SRM (Sakura, Япония), окрашивали гематоксилином и эозином в автомате TISSUE-TEK Prisma (Sakura, Япония) и заключали под пленку в автомате TISSUE-TEK Film (Sakura, Япония).
С целью идентификации ТК использовали окраску препаратов толуидиновым синим (BioVitrum, Россия), согласно рекомендациям фирмы-производителя. Для оценки плоидности гепатоцитов после кислотного гидролиза препараты окрашивали по Фельгену. Высчитывали средний индекс накопления ДНК (ИНДНК) в 25–30 гепатоцитах. Фотографирование препаратов, а также морфометрию гепатоцитов, ТК и плоиднометрию проводили с использованием комплекса визуализации МC-LCD 4K (Россия), микроскопа «Микмед-6» (АО «ЛОМО», Россия) и программы «ВидеоТест-Морфология 5.2.» (ООО «Видеотест», Россия). Подсчет плотности распределения ТК в ткани печени осуществляли в программе UTHSCSA Image Tool 3.0 (разработанной в University of the Texas Health Science Cennter of San Antonio, Texas) в 10 полях зрения (площадь поля зрения составляла 0,12 мм²) при увеличении микроскопа в 400 раз. По морфологическим критериям в процентном отношении подсчитывали недегранулированные (компактные) формы и клетки в состоянии дегрануляции (в том числе в состоянии лизиса). Для определения степени зрелости ТК гистологические срезы окрашивали последовательно 1%-ным раствором альцианового синего на 3%-ной уксусной кислоте (рН 3,2) и раствором 0,5%-ного сафранина в 0,1 N HCl (рН 1,5) и оценивали число незрелых альцианположительных (альциан +), промежуточных, содержащих синие и красные гранулы (альциан-сафранин ++), и зрелых сафранинположительных (сафранин +) форм ТК.
Полученный информационный массив данных обрабатывали при помощи статистического пакета Statistica 12.0 (StatSoft, USA). Различия признавались статистически значимыми при p < 0,05.
Результаты исследования и их обсуждение. При микроскопическом исследовании печеночные дольки имели многоугольную форму, были окружены умеренно выраженной соединительной тканью, границы долек определялись чётко. Гепатоциты выглядели полиморфными: они были округлой, овальной или кубической формы. Средняя площадь цитоплазмы гепатоцитов составила 470,7±13,8, площадь ядра – 33,8±1,7 мкм², ИНДНК - 3,9±0,2 (табл. 1).
ТК в печени интактных крыс располагались преимущественно в портальных трактах около сосудов и желчных протоков. Они обнаруживались, как правило, поодиночке и имели округлую форму (рис. 1).
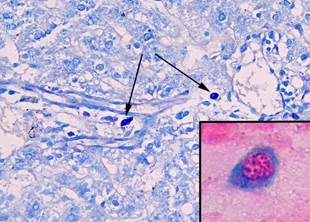
Рис. 1. Тучные клетки в печени интактных животных. При окраске толуидиновым синим тучные клетки небольшого размера с компактным расположением гранул (показаны стрелками), увеличение х 400. В рамке на большом увеличении незрелая тучная клетка (цитоплазма окрашена альциановым синим в синий цвет), фрагмент микрофотографии увеличен с помощью программного обеспечения, окраска альциановым синим – сафранином
Сафранин + ТК при окраске альциановым синим – сафранином обнаружены не были, альциан-сафранин ++ ТК было 27,4 ± 8,5%, а альциан + ТК было 72,6 ± 8,5%. Среднее количество ТК составило 1,2 ± 0,2 в 10 полях зрения, средняя площадь ТК была равна 34,0 ± 0,7 мкм². Количество ТК с компактным расположением гранул составило 83,2 ± 0,3%, а в состоянии дегрануляции было 16,8 ± 0,5% (табл. 2, 3).
Таблица 1
Морфометрические параметры гепатоцитов в зависимости от срока проведения однократной глубокой иммерсионной гипотермии (M±m)
|
Срок эксперимента
|
Морфометрические параметры гепатоцитов |
||
|
Площадь ядра (мкм2) |
Площадь цитоплазмы (мкм2) |
ИНДНК (с) |
|
|
Интактные животные (1) |
33,8±1,7 |
470,7±13,8 |
3,9±0,2 |
|
Сразу после гипотермии (2) |
26,5±1,7 |
364,4±18,9 |
3,2±0,2 |
|
Через 2 дня (3) |
45,0±1,7 |
501,7±40,8 |
5,7±0,2 |
|
Через 7 дней (4) |
100,3±6,3 |
394,2±10,8 |
9,3с±0,2 |
|
Через 14 дней (5) |
67,2±4,2 |
425,3±40,1 |
5,7с±0,2 |
|
Через 30 дней (6) |
63,9±3,5 |
479,4±31,3 |
4,5с±0,2 |
Примечание: Р для площади ядра -Р1-2 = 0,004; Р2-3 = 0,004; Р3-4 = 0,0000001; Р4-5 = 0,0000001; Р5-6 =0,4;
Р для площади цитоплазмы ядра -Р1-2 = 0,0002; Р2-3= 0,0002; Р3-4 = 0,007; Р4-5 = 0,4; Р5-6 =0,3.
Р для ИНДНК -Р для ИНДНК -Р1-2 = 0,01; Р2-3 = 0,0000001; Р3-4 = 0,0000001; Р4-5 = 0,001; Р5-6 =0,0001.
Таблица 2
Характеристика тинкториальных особенностей тучных клеток в зависимости от срока эксперимента (M±m)
|
Срок эксперимента |
Альциан (+) ТК (%) |
Альциан- сафранин ТК (++) (%) |
Сафранин (+) ТК (%) |
|
Интактные животные (1) |
72,6±8,5 |
27,4±8,5 |
- |
|
Сразу после гипотермии (2) |
43,1±12,4 |
24,8±9,7 |
32,1±12, 4 |
|
Через 2 дня (3) |
- |
27,8±8,7 |
72,2±8,7 |
|
Через 7 дней (4) |
7,9±3,7 |
22,6±7,0 |
69,4±8,7 |
|
Через 14 дней (5) |
10,0±4,7 |
22,2±8,1 |
67,8±10,5 |
|
Через 30 дней (6) |
32,7±5,1 |
56,3±5,2 |
10,9±3,8 |
Примечание: Р для альциан + ТК – Р1-2 = 0,05, Р2-4= 0,003; Р4-5= 0,9; Р5-6 =0,02;
Р для альциан-сафранин ++ ТК –Р1-2 = 0,8, Р2-3= 0,8; Р3-4= 0,6; Р4-5= 0,9; Р5-6=0,001;
Р для сафранин + ТК – Р2-3= 0,00001; Р3-4 = 0,0001; Р4-5 = 0,9; Р5-6 =0,01.
Таблица 3
Количество, площадь и состояние тучных клеток в зависимости от срока эксперимента (M±m)
|
Срок эксперимента |
Параметры тучных клеток |
|||
|
Количество ТК |
Количество компактных форм ТК (%) |
Количество ТК в состоянии дегрануляции (%) |
Площадь ТК (мкм2) |
|
|
Интактные животные (1) |
1,2±0,2 |
83,2±0,3 |
16,8±0,5 |
34,0±0,7 |
|
Сразу после гипотермии (2) |
4,0±0,5 |
64,5±5,1 |
35,5±5,1 |
40,2±3,2 |
|
Через 2 дня (3) |
10,2±1,4 |
54,4±0,9 |
45,6±0,9 |
51,5±1,9 |
|
Через 7 дней (4) |
10,8±0,9 |
43,2±1,9 |
56,8±0,19 |
73,3±3,2 |
|
Через 14 дней (5) |
9,0±1,4 |
66,7±3,1 |
33,3±3,1 |
57,9±2,3 |
|
Через 30 дней (6) |
4,5±0,6 |
72,6±4,6 |
27,4±4,6 |
47,9±2,8 |
Примечание: Р для числа ТК – Р1-2 = 0,0001, Р2-3 = 0,0000001; Р3-4 = 0,9; Р4-5 = 0,7; Р5-6 =0,000001;
Р для площади ТК – Р1-2 = 0,04, Р2-3 = 0,0001; Р3-4 = 0,0001; Р4-5 = 0,0001; Р5-6 =0,000001;
Р для числа компактных ТК – Р1-2 = 0,02; Р2-3 = 0,03; Р3-4 = 0,01; Р4-5 =0,001, Р5-6 = 0,03;
Р для числа ТК в состоянии дегрануляции – Р1-2 = 0,01; Р2-3 = 0,01; Р3-4 = 0,001; Р4-5 =0,001, P5-6 = 0,1.
Сразу после проведения однократной глубокой иммерсионной экспериментальной гипотермии (ГИГ) печеночные дольки имели многоугольную форму, границы их были видны нечётко. Гепатоциты были округлой или овальной формы и резко уменьшены в размерах. Средняя площадь цитоплазмы гепатоцитов по сравнению с интактными животными уменьшалась в 1,3 раза (p = 0,0002) и составила 470,7±13,8 мкм², площадь ядра была меньше в 1,3 раза (p = 0,004) (26,5±1,7 мкм²), ИНДНК снижался в 1,2 раза (p = 0,02) (3,2±0,2) (табл. 1). Число альциан + ТК уменьшалось в 1,7 раза (p = 0,05), количество альциан-сафранин ++ ТК снижалось до 24,8±9,7%, и появлялись сафранин + ТК (32,1±12,4%) (рис. 2).
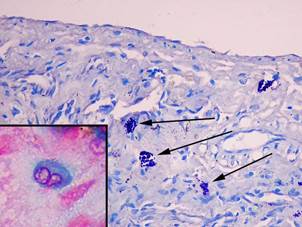
Рис. 2. Тучные клетки в печени крыс сразу после однократной ГИГ. При окраске толуидиновым синим тучные клетки в состоянии дегрануляции (показаны стрелками), увеличение х 400. В рамке незрелая тучная клетка (цитоплазма окрашена альциановым синим в синий цвет), фрагмент микрофотографии увеличен с помощью программного обеспечения, окраска альциановым синим – сафранином
Среднее число ТК в 10 полях зрения в ткани печени возрастало в 3,3 раза (p = 0,0001), средняя площадь ТК увеличивалась до 40,2±3,2 мкм² (p = 0,04), количество клеток в состоянии дегрануляции возрастало в 2,1 раза (p = 0,01), а содержание компактных форм уменьшалось в 1,3 раза (p = 0,02) (табл. 2, 3).
На 2-е сутки после начала эксперимента ткань печени находилась в состоянии умеренно выраженного отека, в гепатоцитах отмечались явления мелкокапельной жировой дистрофии. Гепатоциты имели полиморфную форму, их ядра увеличивались в размерах в 1,7 раза (p = 0,004) до 45,0±1,7 мкм², ИНДНК в ядрах увеличивался в 1,8 раза (р = 0,0000001) до 5,7с±0,2, также в 1,4 раза (p = 0,0002) возрастала площадь цитоплазмы до 501,7±40,8 мкм² (табл. 1).
Альциан + ТК обнаружены не были, число альциан-сафранин ++ ТК возрастало до 27,8±8,7% (p = 0,8), а содержание сафранин + ТК увеличивалось в 2,25 раза (p = 0,00001). Среднее число ТК в 10 полях зрения возрастало в 2,55 раза (p = 0,0000001), средняя площадь их увеличивалась в 1,3 раза, содержание клеток в состоянии дегрануляции было больше в 1,3 раза (p = 0,0001), а число компактных форм уменьшалось в 1,2 раза (p = 0,03) (табл. 2, 3).
На 7-е сутки эксперимента гепатоциты приобретали полиморфную форму и площадь их значительно возрастала. Средняя площадь ядра по сравнению с предыдущим сроком эксперимента была больше в 2,2 раза (p = 0,0000001) (100,3±6,3 мкм²), площадь цитоплазмы на этом сроке эксперимента несколько уменьшалась (в 1,3 раза) (p = 0,02) и составила 394,2±10,8 мкм², а ИНДНК увеличивался до 9,3с±0,2 (p = 0,0000001) (табл. 1).
Число альциан + ТК составило 7,9±3,7%, число альциан-сафранин ++ ТК уменьшалось до 22,6±7,0% (p = 0,6), а содержание сафранин + ТК незначительно снижалось до 69,4±8,7% (p = 0,0001). Среднее число ТК в 10 полях зрения составило 10,8±0,9 (p = 0,9), средняя площадь и число клеток в состоянии дегрануляции возрастало в 1,4 раза (p = 0,0001) и 1,25 раза (p = 0,001) соответственно, а содержание компактных форм ТК уменьшалось в 1,3 раза (p = 0,01) (рис. 3, табл. 1, 2).
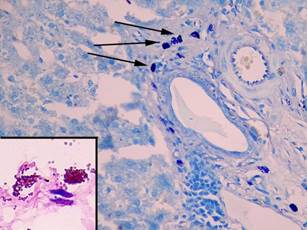
Рис. 3. Тучные клетки в печени крыс на 7-й день эксперимента. При окраске толуидиновым синим большое число тучных клеток в портальном тракте (показаны стрелками), увеличение х 400. В рамке на большом увеличении сафранин+ клетки в состоянии дегрануляции, фрагмент микрофотографии увеличен с помощью программного обеспечения, окраска альциановым синим – сафранином
На 14-е сутки эксперимента по сравнению с 7-м днем отмечали слабо выраженные явления отека, и гепатоциты приобретали округлую форму. Средняя площадь ядра уменьшалась в 1,5 раза (p = 0,0000001) и составила 67,3±4,2 мкм², площадь цитоплазмы гепатоцитов увеличивалась до 425,3±40,1 мкм² (p = 0,4), а ИНДНК уменьшался до 5,7±0,2 (p = 0,001) (табл. 1).
Число альциан + ТК несколько увеличивалось до 10,0±4,7% (p = 0,9), число альциан-сафранин ++ ТК практически не изменялось (22,2±8,1%) (p = 0,9), а содержание сафранин + ТК уменьшалось до 67,8±10,5% (p = 0,9). Среднее число ТК в 10 полях зрения уменьшалось до 9,0±1,4, средняя площадь ТК снижалась в 1,3 раза (p = 0,0001), число клеток в состоянии дегрануляции было меньше в 1,7 раза (p = 0,001), а содержание компактных форм возрастало в 1,5 раза (p = 0,001) (рис. 4, табл. 2, 3).
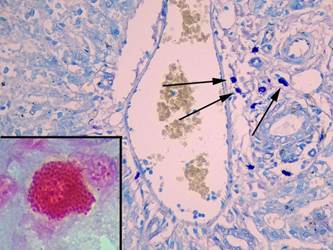
Рис. 4. Тучные клетки в печени крыс на 14-й день эксперимента. При окраске толуидиновым синим видно большое число тучных клеток в портальном тракте (показаны стрелками), увеличение х 400. В рамке на большом увеличении компактная сафранин+клетка, фрагмент микрофотографии увеличен с помощью программного обеспечения, окраска альциановым синим – сафранином
На 30-е сутки по сравнению с 14-м днем размеры ядра и цитоплазмы изменялись незначительно, а гистологическое строение печени было приближено к норме, но ИНДНК в ядрах оставался увеличенным и составил 4,5с±0,2 (р = 0,0001) (табл. 1).
По сравнению с 14-м днем число альциан + ТК возрастало в 3,3 раза (р = 0,02), количество альциан-сафранин ++ ТК увеличивалось в 2,5 раза (р = 0,001), а содержание сафранин + ТК уменьшалось в 6,2 раза (р = 0,01). Среднее число ТК в 10 полях зрения ТК уменьшалось в 2 раза (р = 0,000001) до 9,0±1,4, средняя площадь ТК снижалась в 1,2 раза (p = 0,000001), число клеток в состоянии дегрануляции было меньше в 1,2 раза (р = 0,000001), а содержание компактных форм возрастало до 72,6±4,6% (р = 0,03, табл. 2, 3).
Результаты проведенного исследования показали, что тучноклеточная популяция печени интактных крыс контрольной группы исследования была представлена небольшим количеством преимущественно компактных форм ТК, клетки в состоянии дегрануляции составили 16,8±0,5%, что соответствует данным литературы [16]. При этом преобладали альциан + ТК (72,6±8,5%), которые, по литературным данным, являются незрелыми формами ТК и мигрируют в органы из сосудов микроциркуляции [17]. Возможно, также наличие большого числа незрелых форм связано с низким темпом созревания ТК или низким уровнем факторов, стимулирующих их созревание у крыс.
Гипотермия оказывала повреждающее действие на ткань печени: площадь ядра, цитоплазмы и индекс ДНК гепатоцитов при этом резко уменьшались, что было обусловлено спазмом сосудов, централизацией кровообращения, выключением печени из кровотока и гипоксией. По данным литературы, гипоксия активирует ТК, повышает их миграцию, созревание и дегрануляцию [18]. В нашем исследовании сразу после воздействия гипотермии реакция ТК на холодовой стресс характеризовалась увеличением числа ТК, возрастанием альциан-сафранин ++ форм ТК (что свидетельствует о созревании незрелых ТК и накоплении ими гепарина, который необходим для регенерации клеток печени). Дегрануляция ТК также возрастала.
На 2-й день эксперимента отмечали начало адаптационных компенсаторно-приспособительных процессов в ткани печени: размеры ядер гепатоцитов возрастали в 1,7 раза, а пролиферация клеток, по данным исследования ИНДНК, возрастала в 1,8 раза. На этом этапе эксперимента число сафранин+ ТК было наибольшим, количество альциан-сафранин++ клеток продолжало возрастать, а альциан+ клеток отсутствовали, число клеток в состоянии дегрануляции возрастало.
На 7-й день эксперимента активные адаптивные регенераторные процессы в ткани печени достигали максимума, что соответствует данным литературы, полученным на других моделях [19]. По сравнению с 1-м днем эксперимента площадь ядер гепатоцитов возрастала в 3,8 раза, а ИНДНК увеличивался в 2,9 раза. На этом сроке эксперимента число и площадь ТК были наибольшими, что, по-видимому, было связано с их высокой синтетической активностью. Сафранин+ ТК было 67,8±10,5%, альциан-сафранин++ клеток было 22,6±7,0%, а альциан+ клеток было лишь 7,9±3,7%, возможно, данные ТК являлись незрелыми формами, которые мигрировали в ткань печени из кровяного русла.
Через 14 дней адаптивные процессы стихали, уменьшались размеры ядер гепатоцитов в 1,5 раза и ИНДНК в 1,6 раза. Тинкториальные особенности ТК изменялись незначительно, что свидетельствует об активности ТК на фоне продолжения снижения регенераторных компенсаторно-приспособительных процессов.
На 30-й день эксперимента сафранин+ ТК было лишь 10,9±3,8%, а число альциан-сафранин++ клеток было наибольшим (56,3±5,2%). Нельзя исключить, что данный феномен связан с обратным процессом перехода сафранин+ форм ТК через альциан-сафранин++ формы в альциан+ ТК, которые синтезируют слабо сульфатированные гликозаминогликаны.
Таким образом, выявленные количественные изменения, а также изменения тинкториальных особенностей тучноклеточной популяции после воздействия однократной ГИГ являются следствием регенераторных и адаптивных компенсаторно-приспособительных процессов.
Заключение. Таким образом, выявленные изменения количества и морфологии тучных клеток в печени крыс после однократной глубокой иммерсионной гипотермии заключаются в увеличении как общего количества ТК, так и ТК в состоянии дегрануляции. При этом происходит постепенное замещение альциан+ на сафранин+ ТК. Подобная динамика ТК, скорее всего, является следствием регенераторных и адаптивных компенсаторно-приспособительных процессов.
Conflict of interest
Acknowledgements
Financing
Библиографическая ссылка
Долгатова П.А., Калин Д.А., Бобров И.П., Долгатов А.Ю., Лепилов А.В., Корсиков Н.А., Долгатова Е.С., Лушникова Е.Л., Бакарев М.А. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ КОЛИЧЕСТВА И СОСТОЯНИЯ ТУЧНЫХ КЛЕТОК ПЕЧЕНИ КРЫС ПРИ ГИПОТЕРМИИ // Современные проблемы науки и образования. 2025. № 6. ;URL: https://science-education.ru/en/article/view?id=34330 (дата обращения: 07.07.2026).
DOI: https://doi.org/10.17513/spno.34330