


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
TIME PARAMETERS OF MODELING BLOUNT'S DISEASE IN AN EXPERIMENT

Болезнь Блаунта является одной из сложных патологий опорно-двигательного аппарата у детей [1]. Заболевание представляет собой асимметричное нарушение развития и роста проксимального (эпиметафизарного) отдела большеберцовой кости, приводящее к трехмерной деформации нижней конечности [2, 3]. Лечение консервативными методами в большинстве случаев неэффективно. При прогрессировании патологического процесса и деформации единственным вариантом выступает оперативное вмешательство. Этиологические факторы болезни не определены [4]. В современной литературе представлено ограниченное количество работ по изучению данной патологии. Моделирование болезни Блаунта на животных открывает новые возможности в исследовании этиопатогенетических механизмов рассматриваемого заболевания. В эксперименте на двуногом животном (курах) получены первые результаты по моделированию варусной деформации коленных суставов [5, 6]. Модель создана путем ингибирования экспрессии парного боксового гена 3 (РАХ3) малой интерферирующей рибонуклеиновой кислотой (siРНК) и основана на гипотезе о роли клеток нервного гребня в индуцировании болезни Блаунта. Дифференцировка и миграция клеток нервного гребня регулируются PAX3 геном [7]. К матричной РНК (мРНК) гена РАХ3 через нервную трубку куриного эмбриона вводили siРНК в сроке 53 часов эмбрионального развития на 16-й стадии по классификации Гамбургера и Гамильтона. У цыплят с ингибированием в эмбриогенезе экспрессии РАХ3 гена в постнатальном периоде рентгенологически диагностирована варусная деформация коленных суставов.
Формирование зачатков нижних конечностей у куриного эмбриона происходит на уровне 26–32 пар сомитов [8, 9]. Образование данных пар сомитов представляет собой процесс, протекающий не одномоментно, а происходящий на протяжении нескольких стадий эмбрионального развития и занимающий несколько часов. На 15-й стадии (около 50–55 часов) сформировано 24–27 пар сомитов, на 16-й стадии (приблизительно 51–56 часов) визуализируется 26–28 пар, и 17-я стадия (около 52–64 часов) соответствует 29–32 парам сомитов по классификации Гамбургера–Гамильтона [8]. Поскольку сегментация сомитов происходит от передней к задней части эмбриона и каждая вновь образованная пара сомитов влияет на миграцию клеток нервного гребня в своем окружении [10], ключевым является вопрос выбора времени введения интерферирующей siРНК, направленной к мРНК гена РАХ3. Для индуцирования ортопедической патологии коленного сустава у экспериментального животного важно определить время ингибирования гена РАХ3: siРНК вводить в нервную трубку на этапе начала формирования сомитов, из которых развиваются нижние конечности, или на этапе сформированных нескольких пар сомитов, дающих начало почкам нижних конечностей? В ранее проведенном пилотном эксперименте было выбрано время, соответствующее середине 16-й стадии. В планируемом исследовании представляются необходимыми сравнение нескольких временных параметров и определение оптимального периода для ингибирования экспрессии РАХ3 гена с целью формирования выраженной патологии коленного сустава.
Цель исследования: определить оптимальные временные параметры ингибирования экспрессии парного боксового гена 3 для моделирования болезни Блаунта в эксперименте.
Материал и методы исследования. Проведение экспериментального исследования одобрено Локальным этическим комитетом ФГБУ «ННИИТО им. Я.Л. Цивьяна» Минздрава России (выписка 023/21 из протокола № 009/21 от 06.12.2021 г.).
Эксперимент по моделированию болезни Блаунта у двуногого животного выполняли на яйцах кур породы Росс-308 (ROSS-308 – мясная порода бройлеров). Инкубацию яиц и манипуляции по введению siРНК к мРНК гена PAX3 в нервную трубку куриного эмбриона, закрытию отверстия эмпирического яйца проводили, как описано ранее [6]. Для ингибирования экспрессии гена РАХ3 использовали siРНК, подобранную для проведенного ранее пилотного исследования [6]. В пилотном исследовании было показано, что действие такой siРНК специфично: контрольная siРНК (siScr), не имеющая гомологии с геном PAX3, не оказывала влияния на его экспрессию. Для доставки siРНК в клетки-мишени на основе последовательности siРНК конструировали холестеринсодержащие конъюгаты с конечной концентрацией 50 мМ, хранящиеся при –20°C. Буфер для siРНК: 30 mM HEPES-KOH (pH 7.4), 100 mM ацетат калия, 2 mM ацетат магния.
Экспериментальное исследование включало 3 экспериментальные группы, 3 группы сравнения и контрольную группу. Каждая экспериментальная группа, группы сравнения и контроль состояли из 10 яиц. Количественный состав всего исследования – 70 оплодотворенных яиц. Экспериментальные группы различались временем введения siРНК к мРНК гена PAX3 стеклянным капилляром в нервную трубку куриного эмбриона. В первой экспериментальной группе время введения составило 51–52 часа, во второй – 53–54 часа, в третьей – 55–56 часов эмбрионального развития. Процесс введения осуществляли со стороны каудального отдела эмбриона, располагая стеклянный капилляр под минимально возможным углом к нервной трубке. В первую группу сравнения включены яйца с эмбрионами, которым в 51–52 часа эмбриогенеза был произведен укол стеклянным капилляром без введения вещества. Во второй группе сравнения укол в нервную трубку стеклянным капилляром был выполнен в 53–54 часа, и в третьей группе – в 55–56 часов эмбрионального развития. Процедура укола проводилась аналогично уколу с введением siРНК. Контрольную группу составили интактные яйца. Инкубацию яиц контролировали до конца эмбриогенеза. Для оценки выживаемости птенцов использовали статистический метод относительных показателей. Вылупившихся цыплят содержали в специальных условиях, соответствующих возрасту птицы, обеспечивающих сбалансированное питание на основе комбикормов, обогащенных белково-витаминными-минеральными комплексами и исключающими возможность развития ожирения. В 1 и 3,5 месяца постнатального развития животным из экспериментальных групп 1 и 2, контрольной группы, а также 1-й и 2-й групп сравнения проведена мультиспиральная компьютерная томография (МСКТ) коленных суставов на двухсрезовом компьютерном томографе GE Healthcare Brivo CT325 (США). В 4 месяца курам из тех же групп выполнено обследование нижних конечностей на цифровой рентгенографической системе GE Discovery XR656 (США). Для определения средних значений углов: угла медиального метафизарного «клюва» (MMB) и метафизарно-диафизарного угла (MDA) – использовали метод вычисления средней арифметической величины. При анализе результатов томографической и рентгенологической диагностики руководствовались терминологией, принятой для описательной анатомии опорно-двигательного аппарата птиц [11, с. 24].
Результаты исследования и их обсуждение. На первом этапе исследования была проведена оценка влияния эксперимента на выживаемость птенцов (рис. 1). Выживаемость определяли по способности к вылуплению, она соответствовала проценту вылупившихся из яйца цыплят на 21–22-е сутки инкубации. Оценивали воздействие укола стеклянным капилляром с введением или без введения вещества в нервную трубку куриного эмбриона. Укол осуществляли в разные временные периоды (51–52, 53–54, 55–56 часов) 16–17-й стадий эмбриогенеза по классификации Гамбургера–Гамильтона. Исследование показало, что укол капилляром в нервную трубку с целью введения интерферирующей siРНК (экспериментальная группа 1) или без введения веществ (сравнительная группа 1) на сроке 51–52 часа эмбрионального развития приводит к 80%-ной выживаемости цыплят (рис. 1). При уколе капилляром на сроке 53–54 часа эмбриогенеза из инкубационных яиц вылуплялось 60% и 50% цыплят в экспериментальной группе 2 и в группе сравнения 2 соответственно. Проведение манипуляций по введению вещества или простое физическое воздействие капилляром в период 55–56 часов эмбрионального развития приводили к гибели эмбрионов еще до вылупления в 80–90% случаев. Выживаемость птенцов в 3-й экспериментальной группе составила 10%, а в группе сравнения – 3–20%. Поскольку показатели выживаемости в экспериментальных группах по сравнению с группами сравнения с аналогичным временем укола достоверно не различаются, допустимо сделать вывод, что ключевую роль в выведении птенцов после эксперимента играет время проведения манипуляции, а не введенное вещество. Полученные результаты могут быть связаны с возможной травматизацией эмбриона на более поздних стадиях развития, которые являются критическими. Вероятно, происходит повреждение такого провизорного органа, как амнион. В 55–56 часов эмбрион соответствует 16-й (51–56 часов) – 17-й стадии (52–64 часа) эмбриогенеза [8, 9]. В этот период амнион простирается ниже уровня 18-го сомита и может быть разной длины. Визуально оценить границы амниона во время укола не представляется возможным, поэтому манипуляции стеклянным капилляром с каудальной части эмбриона могли привести к травмированию и гибели последнего.

Рис. 1. Оценка выживаемости цыплят после эксперимента: ЭГ1 – экспериментальная группа 1, ЭГ2 – экспериментальная группа 2, ЭГ3 – экспериментальная группа 3, ГС1 – группа сравнения 1, ГС2 – группа сравнения 2, ГС3 – группа сравнения 3, К – контрольная группа
Животные из экспериментальной группы 3 и из 3-й группы сравнения были исключены из следующего этапа исследования. Проведение эксперимента во временные промежутки 55–56 часов эмбрионального развития для моделирования ортопедической патологии представляется нецелесообразным в связи с низким процентом выживаемости птенцов.
На втором этапе эксперимента оценивали влияние интерферирующей siРНК, направленной к мРНК гена PAX3, на формирование ортопедической патологии коленных суставов у цыпленка в постнатальном периоде. В 1 и 3,5 месяца после вылупления животным из 2 экспериментальных, контрольной и 2 групп сравнения была проведена мультиспиральная компьютерная томография (МСКТ). Исследование показало отсутствие структурных нарушений нижних конечностей у цыплят из контрольной группы, вылупившихся из интактных яиц (рис. 2). Не диагностировано патологических изменений тазобедренных, коленных и голеноплюсневых суставов и у животных из 1-й и 2-й групп сравнения в 1 и 3,5 месяца постнатального развития после физического воздействия на этапе эмбриогенеза. Таким образом, укол стеклянным капилляром без введения вещества в нервную трубку развивающегося куриного эмбриона не приводит к нарушению нормального формирования скелета нижних конечностей.
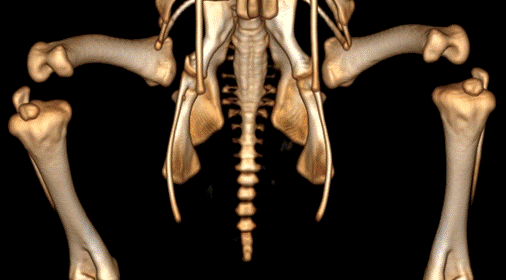
Рис. 2. Трехмерная реконструкция нижних конечностей цыпленка из группы контроля в 1 месяц постнатального развития. Вид спереди
Томографическое исследование нижних конечностей цыплят из экспериментальной группы 1 с введением в нервную трубку куриного эмбриона интерферирующей siРНК во временной промежуток 51–52 часа эмбриогенеза в 1 месяц постнатального развития не выявило структурных нарушений компонентов коленного сустава. Конфигурация бедренной и большеберцовой костей не изменена, контуры ровные и четкие. Малоберцовая кость полностью сформирована. Соотношение костей в коленном суставе правильное. У 50% цыплят данной группы детектирована задержка процесса оссификации большеберцовой кости обеих конечностей (рис. 3а, 3б).
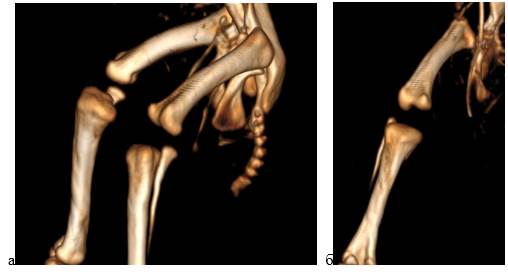
Рис. 3. Трехмерная реконструкция нижних конечностей цыпленка из экспериментальной группы 1 в 1 месяц постнатального развития.
Вид сбоку (а) и спереди (б)
На мультипланарных реконструкциях не визуализируется эпифизарный центр окостенения проксимального отдела большеберцовой кости, наблюдаемый в контроле и у остальных животных экспериментальной группы 1. Сканирование нижних конечностей животных из экспериментальной группы 2 с введением в нервную трубку куриного эмбриона интерферирующей siРНК во временной промежуток 53–54 часа эмбриогенеза не выявило структурных нарушений и задержки оссификации компонентов коленного сустава по сравнению с контролем.
МСКТ-исследование животных из экспериментальной группы 1 в возрасте 3,5 месяца показало, что на трехмерных моделях нижних конечностей отсутствует центр оссификации коленной чашечки правой и левой бедренных костей (рис. 4а). Выявленные изменения обнаружены у 50% цыплят с задержкой остеогенеза, установленной в 1 месяц постнатального развития. Надколенник у этих животных сформирован хрящевой тканью, не визуализирующейся при 3D-реконструкции. Известно, что окостенение рассматриваемого компонента коленного сустава в норме начинается на 12-й неделе после вылупления [11, с.58]. Томографическое исследование нижних конечностей остальных цыплят данной группы свидетельствует о том, что центр окостенения коленной чашечки детектируется и не отличается от аналогичного в группе контроля (рис. 4б). У животных из экспериментальной группы 2 в возрасте 3,5 месяца изображения коленных суставов в сагиттальной, фронтальной и аксиальной проекциях показали отсутствие изменений формы и размеров большеберцовой и бедренной костей по сравнению с контрольной группой. Контуры костей обеих конечностей ровные и четкие. Центр окостенения надколенника визуализируется.

Рис. 4. Трехмерная реконструкция нижних конечностей цыпленка из экспериментальной группы 1 (а) и группы контроля (б) в 3,5 месяца постнатального развития. Вид сбоку
Таким образом, томографическое исследование позволило выявить нарушения развития компонентов коленного сустава в первые месяцы онтогенеза цыпленка, когда рентгенологическое обследование нерезультативно в связи со значительной долей хрящевой ткани. По данным компьютерной томографии можно сделать вывод о замедлении процесса остеогенеза у 50% животных из экспериментальной группы 1. О нарушении нормального развития коленного сустава свидетельствуют задержка оссификации эпифиза проксимального отдела большеберцовой кости в 30 дней постнатального развития и отсутствие очагов окостенения коленной чашечки в возрасте 3,5 месяца. Выявленные изменения не детектированы у остальных цыплят из экспериментальной группы 1 и животных экспериментальной группы 2. Полученные результаты согласуются с данными гистологического исследования о нарушении нормального окостенения в проксимальном отделе большеберцовой кости у детей с болезнью Блаунта [12]. Методом томографии не диагностировано структурных нарушений тазобедренного и голеноплюсневого суставов во всех группах исследования.
Рентгенологическое исследование нижних конечностей цыплят из экспериментальных групп 1 и 2, групп сравнения и группы контроля показало, что в возрасте 4 месяцев постнатального развития все компоненты коленного сустава сформированы. Визуализируются ровные, четкие суставные поверхности бедренной и большеберцовой костей, малоберцовая кость и коленная чашечка. У животных из экспериментальных групп 1 и 2 в проксимальном отделе большеберцовой кости обеих конечностей выявлены расширение и смещение медиального метафиза с формированием выпуклости, похожей на клюв (рис. 5).
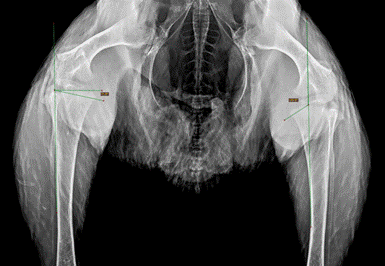
Рис. 5. Рентгенограмма нижних конечностей цыпленка с патологией коленных суставов, 125 дней онтогенеза. Угол MDA 11,8°, угол ММВ 122°
На переднезадних скрининговых рентгенограммах цыплят было произведено определение угла медиального метафизарного «клюва» (MMB) и метафизарно-диафизарного угла (MDA), играющих ключевую роль в диагностике болезни Блаунта [1, 2]. Измерение показало, что средняя величина угла MMB в экспериментальной группе 1 составляет 124° (рис. 5). Среднее значение угла MDA в данной группе равно 13°. Аналогичные результаты получены в экспериментальной группе 2: средний метафизарно-диафизарный угол соответствовал 15°, а величина угла медиального метафизарного клюва – 123°. В группе контроля и группах сравнения 1 и 2 средние значения измеряемых показателей составили: угол MMB 107°, угол MDA 8°. Таким образом, у животных в экспериментальных группах 1 и 2 диагностированы повышенные значения измеряемых углов MDA и MMB по сравнению с животными без введения siРНК в нервную трубку в эмбриональном периоде. Различия в величине углов между экспериментальными группами 1 и 2 отсутствуют.
Рентгенологические данные свидетельствуют о том, что у животных с введением на этапе эмбриогенеза интерферирующей siРНК к гену РАХ3 в постнатальном периоде диагностированы изменения медиальной части проксимального отдела большеберцовой кости, характерные для болезни Блаунта. Полученные результаты по средней величине углов MMB и MDA не отличаются от данных пилотного исследования [6]. Представленные рентгенологические показатели подтверждаются проведенными наблюдением и обследованием экспериментальных цыплят. В 1 месяц постнатального развития у животных в области коленных суставов пальпировались выступающие медиальные мыщелки, тактильно не определяемые у группы контроля. Анализ двигательной активности животных из экспериментальных групп 1 и 2 выявил быструю утомляемость при хождении, невозможность длительного нахождения в положении стоя и прохождения расстояния 1–2 м без остановок и отдыха. О нарушении походки свидетельствовало «переваливание» с ноги на ногу («утиная походка»). Обнаруженные отклонения наблюдались с месячного возраста и были наиболее выражены в 4 месяца. Переломов костей не диагностировано. Хромота и ограничение движений не зафиксированы.
Таким образом, в результате исследования получены данные, позволяющие определить оптимальные временные параметры ингибирования экспрессии РАХ3 гена для моделирования врожденной варусной деформации коленных суставов. Показано, что выживаемость цыплят максимальна после проведения эксперимента в начале 16-й стадии по классификации Гамбургера–Гамильтона, что соответствует 51–52 часам эмбриогенеза. Проведенное томографическое обследование животных в 1 и 3,5 месяца постнатального развития выявило нарушение процесса остеогенеза большеберцовой кости и коленной чашечки у 50% птенцов из экспериментальной группы 1. При рентгенологическом исследовании обнаружено увеличение углов MDA и MMB у цыплят в экспериментальных группах 1 и 2 по сравнению с контролем. Представленные результаты позволяют сделать вывод, что оптимальным временем для проведения эксперимента с целью индуцирования болезни Блаунта у куриц является период 51–52 часа эмбрионального развития (экспериментальная группа 1). Показатели выживаемости в 1-й экспериментальной группе наиболее приближены к контролю, а рентгенологические показатели не отличаются от таковых в экспериментальной группе 2.
Заключение. В рамках гипотезы о роли клеток нервного гребня в индуцировании болезни Блаунта проведено исследование с целью определения оптимальных временных параметров ингибирования экспрессии гена РАХ3 для моделирования врожденной варусной деформации коленных суставов в эксперименте. Полученные данные свидетельствуют о том, что выживаемость цыплят максимальна при введении интерферирующей siРНК в нервную трубку куриного эмбриона в начале 16-й стадии по классификации Гамбургера–Гамильтона. Томографическое исследование в первые месяцы постнатального развития цыпленка выявило нарушение процесса оссификации большеберцовой кости и коленной чашечки у 50% кур с индуцированной патологией коленного сустава. Рентгенологически у экспериментальных животных диагностированы изменения медиальной части проксимального отдела большеберцовой кости, характерные для болезни Блаунта. Исследование продолжается.
Библиографическая ссылка
Строкова Е.Л., Пахомова Н.Ю., Черноловская Е.Л., Жуков Д.В., Кожевников В.В., Гутт А.А., Корыткин А.А. ВРЕМЕННЫЕ ПАРАМЕТРЫ МОДЕЛИРОВАНИЯ БОЛЕЗНИ БЛАУНТА В ЭКСПЕРИМЕНТЕ // Современные проблемы науки и образования. 2025. № 1. ;URL: https://science-education.ru/en/article/view?id=33795 (дата обращения: 03.08.2026).
DOI: https://doi.org/10.17513/spno.33795