


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
HISTOTOPOGRAPHIC AND STRUCTURAL FEATURES OF THYROID FOLLICLES UNDER TRAINING LOADS OF DIFFERENT DURATION

Формирование срочной и долговременной адаптации организма к различным режимам двигательной активности обеспечивается, в том числе, и изменением активности щитовидной железы. Функциональной единицей органа является фолликул [1], характеристики его площади и объема определяются соотношением фолликулярного эпителия и коллоида [2]. Известно, что активность органа прямо пропорциональна относительному объему эпителия и обратно пропорциональна содержанию коллоида [3]. Исследованиями, проведенными нами [4], показано, что в зависимости от величины и продолжительности двигательной активности изменения структурно-функциональных единиц железы имеют мозаичный характер. Поэтому точная оценка морфологических показателей активности фолликулов разных гистотопографических зон обеспечивает исследователя объективной информацией о состоянии органа.
Цель исследования: оценить влияние однократных и многократных тренирующих нагрузок (бег на ленте тредмилла) на структурные и гистотопографические особенности фолликулов щитовидной железы собак.
Материалы и методы исследования
Работа выполнена на собаках-самцах от 1,5 до 2 лет, которые представлены контрольной (16 животных) и двумя экспериментальными группами, получавшими однократные нагрузки тренирующего характера (экспериментальная группа 1) в виде бега на ленте тредмилла при динамическом контроле ЧСС (8 животных, время бега – 27,76±4,67 мин) и многократные тренировки (Многократные 2 – 10 животных, общее количество нагрузок – 55,50±2,45, время бега – 64,02±3,4 мин; Многократные 3 – 11 животных, общее количество нагрузок – 70,54±2,52, время бега – 71,3±2,56 мин).
Исследования на животных проведены в соответствии с приказами Минвуза СССР от 13.11.1984 г. № 742 «Об утверждении правил проведения работ с использованием экспериментальных животных», от 23.01.1985 г. № 48 «О контроле за проведением работ с использованием экспериментальных животных» и положениями европейской Конвенции о защите позвоночных животных, используемых для экспериментов или в иных научных целях (1986г.). Проведение исследования одобрено локальным этическим комитетом от 21.12.2018 г. №313. Исследования проводились на архивном материале.
Взятие материала осуществляли под внутривенным общим комбинированным наркозом (Золетил 50 в дозе 5 мг на 1 кг массы животного, с премедикацией 2%-ным ромитаром внутримышечно в дозе 0,2 мл на 1 кг массы животного) и управляемым внешним дыханием в стандартное время суток – 10–12 часов. Анализу подвергались поперечные срезы парафин-целлоидиновых блоков (фиксатор – 10%-ный нейтральный формалин) толщиной 3 мкм, изготовленные с помощью роторного микротома (ERM-230L) и окрашенные гематоксилин-эозином. Захват кадров (микроскоп «Lеika DMLS» (Leica, Германия), камера «ССВ Camera DIGITAL Kocom» (Kocom, Южная Корея), ув. 400) проводился с учетом топографии всего среза в центральной, промежуточной и периферической зонах относительно его геометрического центра [5]. В программе ImageJ измерялся наибольший (L) и наименьший (B) наружный (ext) и внутренний (int) диаметры (мкм) [6]. Вычисляли: площадь фолликула Sfoll=π×Lext×Вext/4; площадь коллоида Sсoll=π×Lint×Вint/4; площадь эпителия Spar=Sfoll – Scoll. В программе «Statistica StatSoft, Inc., ver.10» проводили сравнение показателей исследуемых групп. Распределение показателей по критерию Колмогорова–Смирнова отличалось от нормального. Проверка статистических гипотез проведена с помощью непараметрического критерия Краскела–Уоллиса, который показал наличие статистически значимых различий между группами при критическом уровне значимости р≤0,05. При дальнейшем попарном сравнении групп использовали U-критерий Манна–Уитни (пересчет критического уровня значимости для устранения эффекта множественных сравнений составил в контроле 0,05/3=0,0167, при сравнении групп – 0,05/6=0,0083). Данные представлены в виде Me (Q1–Q3), мкм2. Квартильные интервалы контрольной группы были критериями деления фолликулов по площади на большие, средние, малые. Определяли процентное соотношение фолликулов разной площади к общему числу, измеренных для группы в целом и отдельных зон. Проводили кластерный анализ, где в качестве объектов выступали площади фолликулов различного диаметра и их представительство в гистотопографических зонах.
Результаты исследования и их обсуждение. Проведенные исследования показали, что площадь фолликулов, коллоида и эпителия при нагрузках уменьшалась и статистически значимо (p<0,02) отличалась от контроля (табл. 1). Статистически значимые различия наблюдались между экспериментальными группами, за исключением значений площади фолликулов (Тренирующие нагрузки и Многократные 3, p=0,05) и площади эпителия (Многократные 2 и Многократные 3, p=0,63).
Изменялись среднегрупповые значения коллоидно-эпителиального отношения: для контрольной группы они составили 77% к 23%, при однократных нагрузках – 75% к 25%, при Многократных 2 – 78% к 22%, при Многократных 3 – 72% к 28%. Под воздействием бега менялось соотношение фолликулов разной площади: в контроле выявлено 8% малых, 58% средних, 33% больших. Нагрузки приводили к увеличению числа малых фолликулов (Однократные тренирующие – 31%, Многократные 2 – 24%, Многократные 3 – 32%) за счет уменьшения количества средних (44, 48, 49%) и больших (в большей степени) фолликулов (25%, 28%, 19% соответственно).
Выявленная динамика свидетельствует об увеличении функциональной активности железы, так как фолликулы меньшего диаметра обеспечивают большую площадь контакта с коллоидом, что создает предпосылки для усиленной резорбции коллоида и продукции тиреоидных гормонов [7, 8]. Исходя из изложенного можно предположить, что ведущим механизмом формирования пула малых фолликулов является их увеличение не только за счет резорбции коллоида средних, но и за счет деления больших фолликулов гемокапиллярами [7, 9], возможен и неофолликулогенез из интерфолликулярного эпителия [10, 11].
Таблица 1
Площадь фолликула, коллоида и паренхимы щитовидной железы у животных контроля и в экспериментальных группах, Me (Q1-Q2) (мкм2)
|
Группы/ Признаки |
Площадь фолликулов |
Площадь коллоида |
Площадь эпителия |
|
Контроль (n=16) 940 фолликулов |
6772 (4534–10653) |
4992 (3183–8136) |
1686 (1233–2356) |
|
Однократные тренирующие (n=8) 845 фолликулов |
4872 #* (2514–9165) |
3443 #* (1580–6781) |
1297 #* (813–2008) |
|
Многократные 2 (n=10) 802 фолликула |
5469 #* (3078–9725) |
3776 #* (2001–7526) |
1502 #* (976–2229) |
|
Многократные 3 (n=11) 1266 фолликулов |
4321 #* (2565–7732) |
2741 #* (1380–5448) |
1506 #* (1006–2237) |
Примечание: при анализе всех полей зрения от контроля # p<0,02, между экспериментальными группами * p<0,01.
Распределение площади фолликулов, коллоида и эпителия относительно геометрического центра в пределах каждой из исследуемых групп варьировало незначимо, в то же время выявлены статистически значимые различия практически всех показателей зон экспериментальных групп относительно контроля (табл. 2).
Таблица 2
Распределение площади фолликулов, коллоида и эпителия щитовидной железы по зонам относительно геометрического центра, Me (Q1–Q3) (мкм2)
|
Зона |
Площадь |
Контроль |
Однократные тренирующие |
Многократные 2 |
Многократные 3 |
|
Центральная |
фолликула |
7191 (4497–10691) |
4940 # (2460–9519) |
6205 (4045–10914) |
4492 # (2761–7656) |
|
коллоида |
5191 (3224–7996) |
3551 # (1524–6813) |
4288 (2621–8420) |
2820 # (1445–5253) |
|
|
эпителия |
1740 (1235–2525) |
1260 # (760–2008) |
1632 (1212 – 2435) |
1547 # (1078–2317) |
|
|
Промежуточная |
фолликула |
6259 (4495–10240) |
4827 # (2514–9190) |
5073 # (2801–9256) |
4316 # (2594–8309) |
|
коллоида |
4572 (3050–7779) |
3535 # (1605–6415) |
3443 # (1701–7233) |
2664 # (1376–5837) |
|
|
эпителия |
1621 (1218–2194) |
1281 # (803–1949) |
1476 # (928–2197) |
1561 (1061–2246) |
|
|
Периферическая |
фолликула |
6967 (4631–11633) |
4862 # (2602–8671) |
5331 # (2959–9169) |
4282 # (2434–7484) |
|
коллоида |
5139 (3325–9006) |
3328 # (1575–6821) |
3717 # (2005–6913) |
2852 # (1389–5346) |
|
|
эпителия |
1675 (1266–2382) |
1365 # (866–2082) |
1435 # (953–2108) |
1418 # (938–2114) |
Примечания: при анализе всех полей зрения статистически значимые различия при сравнении с контролем при # p<0,02.
Коллоидно-эпителиальное отношение гистотопографических зон щитовидной железы каждой из экспериментальных групп было относительно постоянным, отличаясь от показателей в контрольной группе. Как при однократных, так и при многократных тренировках менялось соотношение фолликулов разной площади на срезе органа. Увеличение доли малых (максимально Многократные 3 и минимально Многократные 2) фолликулов экспериментальных групп характерно для всех зон железы. При этом данное увеличение в центральной и промежуточной зонах железы происходит за счет уменьшения количества средних, а в периферической зоне – за счет уменьшения больших фолликулов. Гистотопографические особенности распределения площади фолликулов представлены в виде диаграммы (рисунок).
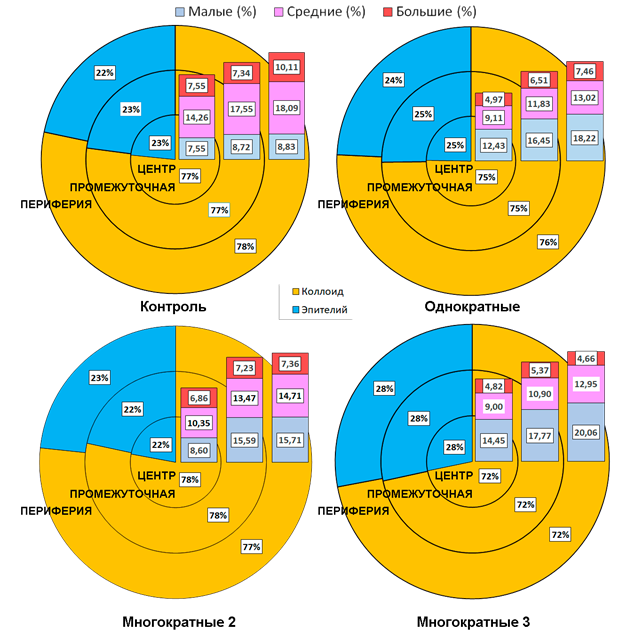
Гистотопография коллоидно-эпителиального отношения и площади фолликулов разных зон щитовидной железы
По данным кластерного анализа в контрольной группе: первый кластер образован связями между периферической и промежуточной зонами, второй – вовлеченностью центральной зоны, а затем последовательным присоединением кластеров со стороны малых, больших и средних фолликулов. Однократные тренировки обусловливают аналогичную реакцию: первый кластер образован связями между периферической и промежуточной зонами, второй – вовлеченностью центральной зоны, а затем последовательным присоединением кластеров со стороны малых, средних и больших фолликулов. Таким образом, при однократных тренирующих нагрузках выявлена первичная реакция со стороны зон железы, а вторичная – со стороны фолликулов разного диаметра. Кластерный анализ группы Многократные 2 выявил первичные изменения со стороны площади фолликулов: первый кластер образован связями между малыми и средними, второй кластер – реакцией со стороны больших фолликулов; вторичные изменения выявлены со стороны гистотопографических зон вначале в центральной, затем в периферической и промежуточной зонах. Для группы Многократные 3 показано, что первичная реакция происходит в зонах органа как в группе контроля и при однократных тренирующих нагрузках: первый кластер образован связями между промежуточной и периферической зонами, второй – присоединением центральной зоны, а вторичная реакция связана с последовательным присоединением малых, средних и больших фолликулов. Таким образом, группа Многократные 2 (общее количество нагрузок 55,50±2,45, время бега 64,02±3,4 мин) и группа Многократные 3 (общее количество нагрузок 70,54±2,52, время бега 71,3±2,56 мин) отличаются, что, вероятно [4], является отражением волнообразности и фазности адаптационного процесса при многократном повторении длительного бега.
Известно, что функциональную единицу щитовидной железы – фолликул – формирует монослой эпителиальных клеток, которые соединены плотными контактами, окружены внеклеточным матриксом, расположены на базальной пластинке, разделены фиброваскулярной стромой, гемо- и лимфокапиллярами, нервами и наполнены коллоидом [1, 7, 8]. Именно в нем происходят биохимические процессы, связанные с синтезом (секрецией и последующей резорбцией) тиреоидных гормонов, производных тиреоглобулина, обеспечивающих адаптацию организма к различным видам воздействий, в том числе к мышечной деятельности [4, 10]. Наши исследования показали, что двигательная активность (бег на ленте тредмилла) приводит к увеличению функциональной активности фолликулов органа.
Морфофункциональные изменения, как правило, затрагивают не единичный фолликул, а их совокупность – структурно-функциональную единицу органа (СФЕ) – ангиофолликулярную единицу, тиреоидный микрорайон или микродольку [9, 12]. Проявлением функционального напряжения СФЕ являются увеличение площади эпителиальных клеток, уменьшение площади коллоида, разделение больших фолликулов на меньшие (в результате чего увеличивается площадь контакта коллоида и эпителия), также возможно формирование новых фолликулов за счет расширения просвета эпителиальных клеток – «фолликулогенез» [9], значимую роль в котором играет апикобазальная поляризация тироцитов [11]. Морфофункциональные изменения в структурно-функциональных единицах [7] происходят асинхронно и, как показано нами, затрагивают разные гистотопографические зоны органа [2].
Заключение. Активация фолликулов щитовидной железы под влиянием бега приводила к статистически значимому уменьшению их площади как при однократных тренирующих нагрузках, так и при многократных повторяющихся тренировках. Изменяется коллоидно-эпителиальное отношение, смещаясь в сторону эпителия (группы Однократные тренирующие и Многократные 3) или коллоида (группа Многократные 2). Исследование фолликулов выявило увеличение доли малых фолликулов за счет уменьшения средних и больших фолликулов во всех гистотопографических зонах железы. В зависимости от параметров двигательной активности (продолжительности и числа тренировок) выявлено два способа реагирования структурно-функциональных единиц железы, обусловленных, вероятно, волнообразностью и фазностью адаптационного процесса. Первичная реакция происходит либо за счет изменения числа фолликулов разной площади (большие, средние, малые), либо за счет изменения пространственной организации гистотопографических зон органа (центральной, промежуточной, периферической).
Библиографическая ссылка
Безденежных А.В., Колодежная С.С., Баврина А.П., Мирошниченко В.В. ГИСТОТОПОГРАФИЧЕСКИЕ И СТРУКТУРНЫЕ ОСОБЕННОСТИ ФОЛЛИКУЛОВ ЩИТОВИДНОЙ ЖЕЛЕЗЫ ПРИ ТРЕНИРУЮЩИХ НАГРУЗКАХ РАЗЛИЧНОЙ ПРОДОЛЖИТЕЛЬНОСТИ // Современные проблемы науки и образования. 2022. № 4. ;URL: https://science-education.ru/en/article/view?id=31911 (дата обращения: 30.06.2026).
DOI: https://doi.org/10.17513/spno.31911