


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
ION-SECRETING FUNCTION OF KIDNEYS IN EXPERIMENTAL ENDOGENOUS HYPERPROLACTINEMIA

Основная роль пролактина, синтезирующегося лактотрофными клетками передней доли гипофиза, заключается в регуляции лактации. Сигнализация пролактина происходит через уникальный трансмембранный рецептор, структура которого в настоящее время выяснена и аналогична структуре многих фундаментальных рецепторов семейства гемопоэтических цитокинов. Рецептор пролактина экспрессируется в широком спектре тканей, и все большее число биологических процессов продолжает приписываться пролактину [1]. Изоформы пролактина выполняют независимые биологические функции, а открытие многочисленных экстрапитуитарных участков секреции пролактина также увеличивает диапазон известных функций гормона, которые влияют на репродуктивную, метаболическую, осмо- и иммунорегулирующие функции, а также на поведение человека [2].
Дофамин, который секретируется гипоталамическими нейронами, является основным ингибирующим регулятором секреции пролактина. Пролактин сам стимулирует секрецию дофамина, тем самым оказывая отрицательную обратную регуляцию на его собственное высвобождение. Пролактин оказывает автокринную/паракринную обратную связь на лактотрофные клетки in vivo, которая может быть полностью компенсирована интактной гипоталамической системой обратной связи, и этот процесс осуществляется двумя различными механизмами: во-первых, опосредованно путем активации гипоталамических дофаминовых нейронов и, во-вторых, непосредственно в гипофизе независимо от дофамина [3].
В физиологических условиях около 25% пролактина выводится с мочой, поэтому при хронической почечной недостаточности у 70% пациенток отмечается повышение уровня пролактина в крови. Особенно часто, до 90%, гиперпролактинемия встречается у больных с терминальной стадией и находящихся на гемодиализе. Эффективное лечение хронической почечной недостаточности сопровождается снижением гиперпролактинемии. Изучение физиологических механизмов влияния пролактина на функции почек в норме и при их патологии, несмотря на имеющиеся клинические и экспериментальные данные, остается актуальным.
Материал и методы исследования
Для выяснения поставленной цели опыты проводили на 50 половозрелых 6–8- месячных лабораторных крысах-самцах линии Вистар массой от 170 до 220 г (190±17 г). Вначале у всех 50 крыс собирали мочу. Полученные результаты отражены в таблицах «Интактный фон (контроль)». Затем у 10 крыс, находящихся в состоянии обезболивания и обездвиживания, вызванного внутрибрюшинным введением анестетика общего действия золетила в дозе 0,1мл/100г массы тела, из сердца набирали кровь в шприц, куда заранее добавили 5–6 капель гепарина (Россия, 5000 МЕ/мл 5,0 мл). Полученные результаты содержания кальция, натрия и калия в плазме крови отражены на рисунке 1 как «Интактный фон». Для выяснения влияния однократного введения нейролептика галоперидола (Венгрия, Гедеон-Рихтер, 0,5% 1,0 мл) оставшимся 40 крысам вводили его в/м в дозе 0,2мг/100г массы тела и на следующий день собирали мочу, после чего у 5 крыс из сердца брали кровь (полученные данные отражены в таблицах и рисунках как «Однократное введение»). Далее мы начали создавать модель эндогенной гиперпролактинемии [4], для чего оставшимся 35 крысам в течение 1 недели вводили галоперидол в указанной дозе и на 8-й день в спонтанном состоянии собирали мочу, а затем у 5 крыс, также в состоянии обезболивания, получали кровь из сердца (данные отражены в таблицах и рисунках «Недельное введение»). Двух-, трех- и четырехнедельное введение нейролептика со сбором мочи на 15-й, 22-й и 28-й дни уже проводили на 30, 25 и 20 крысах, и каждый раз по 5 животных выводили из опытов из-за взятия крови из сердца.
Мочу у крыс собирали в спонтанном состоянии, для чего помещали их на 6 ч в специальные устройства (мочесборники), в которых вся выделившаяся моча в чистом виде поступала в стаканчик. В собранной моче и плазме крови определяли содержание эндогенного креатинина (методом Поппера) и кальция (с эриохромом синим) спектрофотометрически (Unico 2800), а натрия и калия – пламенной фотометрией (ПАЖ-2) [5]. Расчетными показателями были спонтанный диурез, клубочковая фильтрация, канальцевая реабсорбция воды, экскреция кальция, натрия и калия, их фильтрационные заряды, канальцевая реабсорбция и экскретируемые фракции (для кальция и натрия).
По лученные ре зультаты подвергали ст атистической об работке с получением средних значений и отклонений (пр ограмма «G raphPad Pr izm 8.41»), а затем использовали па раметрический ме тод ср авнения ср едних ве личин по классической методике с оценкой статистической значимости по t- критерию Ст ьюдента при р˂0,05.
Разрешение на проведение экспериментов и исследований на животных по изучению функции почек при гиперпролактинемии было выдано этическим комитетом Северо-Осетинской государственной медицинской академии 30 ноября 2017 г., протокол № 7.10.
Работа с животными, постановка оп ытов, утилизация проводились согласно пр иказу «Об ут верждении Пр авил на длежащей ла бораторной пр актики» (МЗ РФ № 199н от 01.04.2016 г.) и Межгосударственному стандарту «Руководство по содержанию и уходу за лабораторными животными» от 01.04.2016 г.
Результаты исследования и их обсуждение
Как мы отмечали ранее, однократное и недельное введение крысам галоперидола со сбором мочи на следующий день не оказало никакого влияния на спонтанный диурез и основные процессы мочеобразования, но начиная с конца второй недели и далее отмечалось статистически значимое снижение диуреза в результате усиления канальцевой реабсорбции воды [6]. Что касается ионовыделительной функции почек, то однократное введение галоперидола и проведение исследований на следующий день в спонтанном состоянии не оказывают влияния на содержание в плазме крови кальция, натрия и калия (рис. 1).
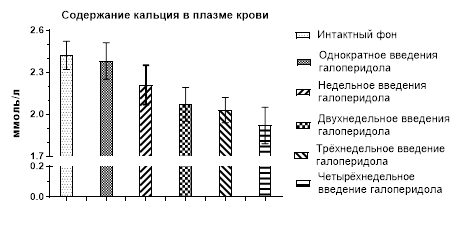
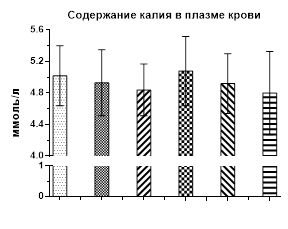
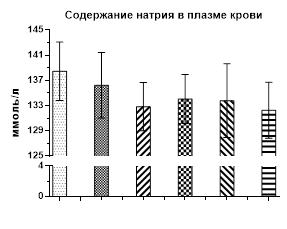
Рис. 1. Содержание кальция, натрия и калия в плазме крови крыс после однократного и 1, 2, 3 и 4 недель введения галоперидола
При недельном введении галоперидола произошло лишь незначительное снижение концентрации кальция в крови, но с продолжением введения препарата еще в течение 1 недели отличие от контроля (2,42±0,10 ммоль/л) приобрело достоверное значение (2,07±0,12ммоль/л; р˂0,05). Это снижение сохранилось и даже несколько усилилось при дальнейшем, еще в течение 2 недель, введении галоперидола (рис. 1). Таким образом, гипокальциемический эффект гиперпролактинемии проявляется только после 2 недель введения галоперидола.
Содержание натрия в крови после недельного введения галоперидола лишь немного снизилось (132,84±3,84 ммоль/л) по отношению к уровню контроля (138,46±4,64 ммоль/л). При продолжении введения нейролептика еще в течение 3 недель уровень натрия в плазме примерно соответствовал контрольному значению (рис. 1).
Введение галоперидола также не оказало влияния на содержание калия в плазме крови на всем протяжении постановки опытов, а отмеченные колебания от контрольных значений были в пределах статистических погрешностей (рис. 1).
Таким образом, создаваемая со второй недели и далее эндогенная гиперпролактинемия, вызванная введением крысам галоперидола, не оказывает влияния на уровень натрия и калия в плазме крови у крыс, но вызывает гипокальциемию.
При выяснении вопроса о том, как однократное введение галоперидола влияет на обработку этих ионов в почках, т.е. на их экскрецию, фильтрационные заряды и обратное всасывание в канальцах почек, были получены результаты, свидетельствующие об отсутствии каких-либо изменений в показателях обмена кальция в почках, отличающихся от таковых интактных крыс контрольной группы (табл. 1).
Таблица 1
Показатели обмена кальция в почках после введения галоперидола
|
Условия опыта |
Стат. показатели |
Экскреция |
Фильтрационный заряд |
Канальцевая реабсорбция |
Экскретируемая фракция |
|
мкмоль/час/100г |
% |
||||
|
Интактный фон (контроль), n=50 |
M±m |
0,68±0,05 |
26,73±1,93 |
97,47±0,19 |
2,53±0,19 |
|
Однократное введение, n=40 |
M±m р |
0,70±0,08 * |
25,45±2,06 * |
97,26±0,17 * |
2,74±0,17 * |
|
Недельное введение, n=35 |
M±m р |
0,84±0,07 * |
28,44±1,85 * |
97,05±0,24 * |
2,95±0,24 * |
|
Двухнедельное введение, n=30 |
M±m р |
1,17±0,09 ˂ 0,001 |
27,38±2,34 * |
95,75±0,18 ˂ 0,001 |
4,25±0,18 ˂ 0,001 |
|
Трехнедельное введение, n=25 |
M±m р |
1,25±0,11 ˂ 0,001 |
29,12±1,88 * |
95,72±0,29 ˂ 0,001 |
4,28±0,29 ˂ 0,001 |
|
Четырехнедельное введение, n=20 |
M±m р |
1,24±0,12 ˂ 0,001 |
29,78±2,25 * |
95,83±0,31 ˂ 0,001 |
4,17±0,31 ˂ 0,001 |
|
Примечание: р – статистически значимые отличия от данных интактных крыс; * – отсутствие статистически значимых отличий. |
|||||
Недельный прием нейролептика лишь незначительно повысил экскрецию кальция, его фильтрационный заряд и экскретируемую фракцию. Но после двухнедельного введения экскреция кальция с контрольного уровня в 0,68±0,05 мкмоль/час/100г статистически значимо (р˂0,001) повысилась до 1,17±0,09 мкмоль/час/100г. При этом фильтрационный заряд не изменился, но снизилась канальцевая реабсорбция (р˂0,001), что и вызвало повышение экскретируемой фракции кальция с 2,53±0,19% до 4,25±0,18% и, как следствие, повышение экскреции кальция.
Проведение исследований после трех- и четырехнедельного введения галоперидола незначительно усилило выведение кальция с мочой и также за счет ослабления его относительной канальцевой реабсорбции, которая способствует повышению экскретируемой фракции иона. Таким образом, причиной выявленного кальцийуреза в условиях эндогенной гиперпролактинемии при неизменности загрузки иона в клубочках нефронов было ослабление относительной канальцевой реабсорбции и, соответственно, повышение экскретируемой фракции.
Очевидно, что создаваемая галоперидолом эндогенная гиперпролактинемия, приводящая к повышению выведения кальция с мочой и снижению его концентрации в плазме крови, должна воздействовать на ремоделирование костной ткани с превалированием резорбции над активностью ее формирования [7].
Данные литературы о влиянии галоперидола на обмен натрия различны: от отсутствия влияния, задержки натрия в организме до усиления его экскреции [8, 9, 10]. В наших исследованиях однократное введение галоперидола не оказало влияния на обмен натрия в почках и ни один из показателей не изменился (табл. 2). Недельное введение, хоть и вызвало незначительно повышение фильтрационного заряда натрия с контрольного уровня в 1937±142 мкмоль/час/100г до 2165±159 мкмоль/час/100г, но так как одновременно немного усилилась канальцевая реабсорбция, то экскреция иона снизилась с 8,34±0,64 мкмоль/час/100г до 7,55±0,73 мкмоль/час/100г.
Таблица 2
Показатели обмена натрия в почках после введения галоперидола
|
Условия опыта |
Стат. показатели |
Экскреция |
Фильтрационный заряд |
Канальцевая реабсорбция |
Экскретируемая фракция |
|
мкмоль/час/100г |
% |
||||
|
Интактный фон (контроль), n=50 |
M±m |
8,34±0,64 |
1937±142 |
99,56±0,05 |
0,44±0,05 |
|
Однократное введение, n=40 |
M±m р |
8,42±0,73 * |
1846±137 * |
99,54±0,07 * |
0,46±0,07 * |
|
Недельное введение, n=35 |
M±m р |
7,55±0,73 * |
2165±159 * |
99,65±0,06 * |
0,35±0,06 * |
|
Двухнедельное введение, n=30 |
M±m р |
5,94±0,31 ˂0,01 |
2246±167 * |
99,73±0,05 ˂ 0,05 |
0,27±0,05 ˂ 0,05 |
|
Трехнедельное введение, n=25 |
M±m р |
5,66±0,42 ˂ 0,01 |
2419±194 * |
99,76±0,05 ˂0,02 |
0,24±0,05 ˂ 0,02 |
|
Четырехнедельное введение, n=20 |
M±m р |
5,71±0,50 ˂ 0,01 |
2520±213 * |
99,77±0,06 ˂ 0,02 |
0,23±0,06 ˂ 0,02 |
|
Примечание: р – статистически значимые отличия к данным интактных крыс; * – отсутствие статистически значимых отличий. |
|||||
Двухнедельное введение галоперидола сопровождалось снижением выделения натрия с мочой (р˂0,01), в основе которого лежало усиление канальцевой реабсорбции иона и соответственно уменьшение его экскретируемой фракции. При продолжении введения галоперидола еще в течение 2 недель экскреция натрия оставалась сниженной по той же причине (табл. 2), и, если натрийурез у интактных крыс принять за 100%, то после недельного введения галоперидола произошло снижение на 9,5%, а после 2, 3 и 4 недель – на 28,8% (р˂0,05), 32,2% (р˂0,01) и 31,6% (р˂0,01) соответственно.
Хотя мы и показали, как меняется обмен натрия в почках, однако механизмы, вызывающие эти изменения, в частности его канальцевая реабсорбция, могут быть обусловлены другими причинами. И одной из них является альдостерон, который усиливает реабсорбцию натрия в канальцах почек, уменьшая тем самым его экскрецию с мочой, и одновременно усиливает секрецию калия и его содержание в моче [11]. В доступной научной литературе имеются данные о том, что повышение содержания пролактина вызывает увеличение секреции альдостерона за счет трех возможных механизмов: первый – увеличение продукции цАМФ, второй – активация Са++-каналов L- и Т-типа, третий – активация 21-гидроксилазы, которая участвует в синтезе альдостерона и кортизола, а также альдостеронсинтазы, катализирующей последнюю стадию синтеза альдостерона в клетках клубочковой зоны [12]. Также есть данные и о том, что при гиперпролактинемии в модели холестаза беременных крыс отмечается снижение активности Na+/K+-АТФазы в мозговом веществе почек, где происходит активная реабсорбция натрия [13].
Результаты изучения обмена калия – основного внутриклеточного катиона – после введения крысам галоперидола представлены ниже. Содержание иона в плазме крови на всем протяжении эксперимента не изменилось, а обработка на уровне почек выглядела следующим образом. Однократное введение галоперидола не вызвало каких-либо изменений (рис. 2), но уже через неделю экскреция калия (5,15±0,42 мкмоль/час/100г), став в 1,4 раза больше, приобрела статистически значимое отличие (р˂0,02) по сравнению с контролем (3,66±0,28 мкмоль/час/100г), но фильтрационный заряд оставался без изменений. Продолжение введения нейролептика способствовало усилению выделения калия с мочой (р˂0,001), которое стало в 1,72 раза больше контрольного уровня в конце второй недели, в 2,15 раза – в конце третьей и в 2,12 раза – при завершении опытов. Загрузка клубочков калием хоть и становилась больше, но это не имело статистически значимого отличия (рис. 2).
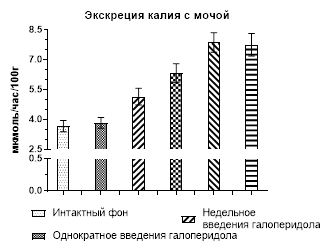
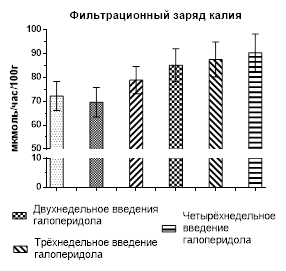
Рис. 2. Экскреция и фильтрационный заряд калия после введения галоперидола
Однако говорить о причине калийуреза можно только в предположении из-за способности калия не только реабсорбироваться в канальцах почек, но и секретироваться, т.е. в этом случае может не только уменьшаться обратное всасывание иона, но и усиливаться его секреция, к чему мы склоняемся в большей степени, так как отмеченное выше снижение экскреции натрия могло быть вызвано повышенным синтезом альдостерона, способного также усиливать секрецию калия.
Заключение
Как однократное, так и недельное введение галоперидола не оказывает влияния на содержание в плазме крови и выделение с мочой кальция, натрия и калия, а двухнедельное введение вызывает снижение содержания кальция в крови и повышение его выделения с мочой из-за ослабления канальцевой реабсорбции иона, уменьшение экскреции натрия вследствие усиления его обратного всасывания в канальцах почек и усиление калийуреза, очевидно, за счет повышения секреции калия. Отмеченные изменения сохраняются и при дальнейшем введении крысам галоперидола в конце третьей и четвертой недель.
Библиографическая ссылка
Джиоев И.Г., Кочиева Д.М., Берёзова Д.Т., Ремизов О.В., Агамирзаев А.А. ИОНОВЫДЕЛИТЕЛЬНАЯ ФУНКЦИЯ ПОЧЕК ПРИ ЭКСПЕРИМЕТАЛЬНОЙ ЭНДОГЕННОЙ ГИПЕРПРОЛАКТИНЕМИИ // Современные проблемы науки и образования. 2021. № 5. ;URL: https://science-education.ru/en/article/view?id=31162 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/spno.31162