


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE MAST CELL INFILTRATION OF THE RATS LUNGS AFTER HYPOTHERMIA

По современным представлениям тучные клетки (ТК) являются ключевым звеном воздействия на метаболические процессы организма. Они играют важную роль при многих физиологических и патологических состояниях, и функции их очень разнообразны. Так, цитокины ТК активируют неоангиогенез и могут стимулировать опухолевый рост [1–3]. Выявлено участие тучноклеточной популяции при стрессовых состояниях [4]. Популяции ТК являются неотъемлемым компонентом микроокружения паренхиматозных компонентов большинства органов, и при их повреждении за счет выделения большого спектра биологически активных веществ мастоциты участвуют в процессах регенерации и компенсаторно-приспособительных процессах [5–7]. Морфофункциональная активность ТК также может стимулировать гипоксия. Так, у крыс при острой массивной кровопотере реакция ТК различных тканей носит тотальный характер и проявляется возрастанием миграции и массивной дегрануляцией [8].
Но эти полифункциональные клетки не только играют значительную роль при различных физиологических и патологических состояниях организма, также они участвуют в его адаптации к действию экстремальных факторов, что позволяет некоторым авторам высказать гипотезу о возможности образования ими единой, самостоятельной регуляторной системы. Перераспределение подвижных ТК между тканями при стрессе, повреждении или гипоксии можно рассматривать в качестве важного компонента адаптационного синдрома [9].
В легких ТК имеют большое значение ввиду многообразных физиологических возможностей и обширных функциональных взаимодействий с клеточным окружением, а также в силу наличия прямых и обратных взаимосвязей с нервной, эндокринной и иммунной системами. Так, в частности, при массивном экзоцитозе гранул они выделяют большое количество гистамина, который вызывает спазм гладкой мускулатуры, в основном мелких бронхов, гиперсекрецию бокаловидных клеток и желез подслизистого слоя трахеобронхиальной системы, повышает проницаемость капилляров межальвеолярных перегородок (в результате чего возникают явления отека); они активирует фибробласты и макрофаги, вызывают миграцию лимфоцитов и нейтрофилов в легкие [10].
В то же время, несмотря на интенсивное изучение морфологии ТК в дыхательной системе в физиологических условиях и при патологии, их морфофункциональные характеристики при действии на легкие экстремальных факторов (и гипотермии в том числе) изучены недостаточно.
Целью исследования являлось изучение влияния одноразовой глубокой иммерсионной гипотермии на морфофункциональную активность тучноклеточной популяции легких крыс Вистар в эксперименте.
Материалы и методы исследования
Исследование было выполнено на 25 самцах крыс линии Вистар массой тела 200–240 граммов. Гипотермию моделировали путем помещения животных, находящихся в индивидуальных клетках, в воду температурой 5°С при температуре окружающего воздуха 7°С. Критерием прекращения воздействия служило достижение животными ректальной температуры 20–25°С, что соответствовало глубокой степени гипотермии. Время экспозиции было индивидуальным и в среднем составляло 40±5 мин. В ходе эксперимента крысы были разделены на 4 группы. Животные 1-й группы (n=5) выводились из эксперимента декапитацией сразу после прекращения охлаждения, животные 2-й группы (n=5) – через 2 суток, животные 3-й группы (n=5) – через 7 суток и животные 4-й группы (n=5) – через 14 суток. Контролем служили 5 животных, которых помещали в индивидуальных клетках в воду температурой 30°С при температуре окружающего воздуха 22–25°С. Время экспозиции соответствовало времени охлаждения животных опытной группы.
Для гистологического исследования образцы легких фиксировали в 10%-ном нейтральном формалине в течение 24–48 часов, затем материал проводили в автомате TISSUE-TEK VIPTM6 (Sakkura, Nagano Japan) и заливали в парафин Histomix (станция парафиновой заливки TISSUE-TEK TEC 5 (Sakkura, Nagano Japan). Срезы толщиной 5–7 мкм изготавливали на роторном микротоме Accu-Cut SRM (Sakkura, Китай), окрашивали гематоксилином и эозином в автомате TISSUE-TEK Prisma (Sakkura, Nagano Japan) и заключали под пленку в автомате TISSUE-TEK Film (Sakkura, Nagano Japan). Окрашивание ТК осуществляли с помощью набора «Толуидиновый синий для окраски тучных клеток» фирмы «Biovitrum» (Санкт-Петербург). Фотографировали препараты при помощи микроскопа Nikon Eclipse E200 (Китай) с цифровой видеокамерой VIDI CAM (Россия) при увеличении х 400. В программе Image Tool 3.0. высчитывали среднюю плотность распределения ТК в 5 полях зрения при увеличении микроскопа х 400. Площадь поля зрения микроскопа составляла 0,365 мм2. Оценивали индекс дегрануляции ТК (ИДТК) (процентное соотношение клеток в состоянии дегрануляции к общему числу анализируемых ТК). Морфометрию ТК проводили с помощью лицензионной морфометрической программы «ВидеоТест – Морфология 5.2». Статистическую обработку полученных данных осуществляли при помощи статистического пакета Statistica 6.0.
Результаты исследования и их обсуждение
При окраске толуидиновым синим в легких контрольной группы животных ТК были видны вокруг бронхиол, бронхов, сосудов и в плевре. Они имели округлую форму, небольшой размер и располагались поодиночке. Плотность распределения ТК в легких интактных животных составила в среднем 2,0±0,5 в поле зрения при увеличении х 400. Площадь клеток составила в среднем 82,5±3,6 мкм2. Число ТК компактных форм ТК было равно 87%±10,1, в состоянии дегрануляции было 23%±10,1.
Сразу после воздействия однократной иммерсионной глубокой гипотермии в ткани легких экспериментальных животных ТК определяли в перибронхиальной ткани. Они были крупными, бледно-фиолетового цвета, округлой или овальной формы (рис. 1).
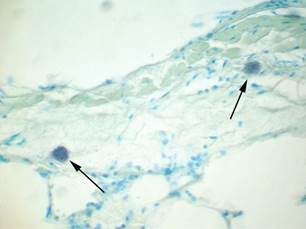
Рис. 1. Легкое крысы. Непосредственно после проведения глубокой гипертермии в перибронхиальной ткани располагается небольшое количество крупных ТК в состоянии гранулоцитолиза (показаны стрелками). Окраска толуидиновым синим. Ув. х400
Плотность распределения ТК сразу после гипотермии в среднем составила 2,7±0,8 в поле зрения, в среднем площадь клеток составила 184,5±14,9 мкм2. Большая часть ТК находилась в состоянии гранулоцитолиза, опустошения в результате секреции гранул. Число ТК в состоянии дегрануляции составило 75%±17,1. Форм ТК, в которых гранулы содержались компактно, было 25%±17,1 (таблица).
Количественная и морфометрическая характеристика тучных клеток легких крыс в зависимости от срока проведения экспериментальной однократной глубокой иммерсионной гипотермии
|
Параметры тучноклеточной инфильтрации |
Срок эксперимента |
|||
|
Сразу после гипотермии (1) |
Через 2 дня (2) |
Через 7 дней (3) |
Через 14 дней (4) |
|
|
Число ТК |
2,5±0,8 |
6,6±0,5 |
8,0±1,0 |
4,3±0,3 |
|
Число компактных ТК (%) |
25,0±17,1 |
52,0±8,9 |
76,7±8,6 |
70,7±12,2 |
|
Число дегранулирующих ТК (%) |
75,0±17,1 |
48,0±8,9 |
23,3±8,6 |
29,3±12,2 |
|
Площадь ТК (мкм2) |
184,5±14,9 |
84,2±3,5 |
107,9±7,3 |
90,8±6,2 |
Примечание: для числа ТК P1-2, P1-3, P1-4 < 0,01. P2-3, P2-4 < 0,04. P3-4 < 0,001. Для числа компактных ТК P1-2, P1-3, P1-4 < 0,001. P2-3 < 0,01. Для числа дегранулирующих ТК P1-2, P1-3, P1-4 < 0,03. P2-3 < 0,04. Для площади ТК P1-2, P1-3, P1-4 < 0,000005. P2-3 < 0,0004.
На 2-е сутки эксперимента в ткани легких ТК располагались группами в стенках межальвеолярных перегородок (рис. 2). ТК имели небольшие размеры и были округлой или овальной формы. Плотность распределения ТК на данном сроке исследования составила 6,6±0,5 в поле зрения, их площадь в среднем составила 84,2±3,5 мкм2. Число компактных форм ТК составило 52,0%±8,9, дегранулирующих форм было 48,0%±8,9 (таблица).
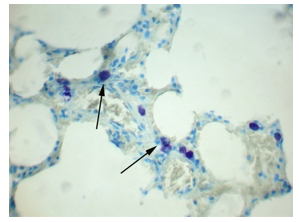
Рис. 2. Легкое крысы. Через 2 суток после гипотермии ТК расположены в капиллярах межальвеолярных перегородок (показаны стрелками). Окраска толуидиновым синим. Ув. х400
На 7-е сутки эксперимента отмечали передвижение ТК к бронхам и бронхиолам, где они располагались в перибронхиальной соединительной ткани, число ТК в межальвеолярных перегородках значительно уменьшалось (рис. 3).
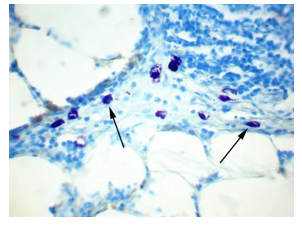
Рис. 3. Легкое крысы. Через 7 суток после гипотермии ТК мигрировали из межальвеолярных перегородок в перибронхиальную ткань (показаны стрелками). Окраска толуидиновым синим. Ув. х400
На данном сроке исследования по своим морфологическим характеристикам ТК отличались от клеток предыдущего срока исследования: они были более интенсивно окрашены, крупных размеров, преобладали клетки удлиненной и неправильной формы. Плотность распределения ТК на данном сроке исследования составила 8,0±1,0 в поле зрения, их средняя площадь увеличилась до 107,9±7,35 мкм2. Компактных форм ТК было 76,0%±8,6, ТК в состоянии дегрануляции – 23,3%±8,6 (таблица).
На 14-й день эксперимента ТК располагались преимущественно поодиночке вокруг бронхиол и бронхов. По сравнению с предыдущим сроком эксперимента ТК были малых размеров, преобладали клетки округлой формы (рис. 4).
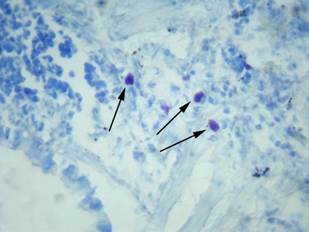
Рис. 4. Легкое крысы. Через 14 суток после гипотермии ТК уменьшались в размерах и были расположены преимущественно в перибронхиальной ткани (показаны стрелками). Окраска толуидиновым синим. Ув. х400
Плотность распределения ТК на данном сроке исследования составила 4,3±0,3 в поле зрения, площадь клеток в среднем была равна 90,8±6,2 мкм2. Количество компактных форм ТК составило 70,7%±13,2, в состоянии дегрануляции было 29,3%±12,2 (таблица).
Обсуждение
Таким образом, полученные данные свидетельствуют о том, что однократная глубокая иммерсионная гипотермия оказывала значительное влияние на морфофункциональную активность ТК легких крыс. Непосредственно сразу после воздействия гипотермии большинство ТК находились в состоянии гранулолизиса или тотальной дегрануляции, что может говорить об истощении системы данных клеток при воздействии холодового стресса. В постгипотермический период, на 2-й день после гипотермии, величина популяции ТК возрастала, большинство клеток было небольшого размера, они локализовались в капиллярах межальвеолярных перегородок, что может свидетельствовать о пополнении резерва популяции ТК за счет миграции в легкие их молодых форм из кровяного русла. На 7-й день эксперимента из альвеол ТК перемещались в перибронхиальную соединительную ткань, число дегранулирующих форм при этом уменьшалось, и на 14-й день содержание таких форм было наименьшим, а количество ТК было близко к норме.
Обнаруженная динамика изменений морфофункциональной активности тучноклеточной популяции при однократной глубокой иммерсионной гипотермии, по нашему мнению, была обусловлена адаптационными процессами. Как известно, в развитии адаптивных реакций выделяют два этапа: 1) этап срочной, но не совершенной адаптации; 2) этап долговременной устойчивой адаптации. Этап срочный адаптивной реакции развивается непосредственно после начала воздействия раздражителя и реализуется на основе готовых, ранее сформированных биологических механизмов. На этом этапе адаптационный эффект обеспечивается не в полной мере, а реакции организма осуществляются на предельных физиологических возможностях. Важнейшим компонентом срочной адаптивной реакции является стресс-реакция со всеми ее проявлениями: повреждением и предельной мобилизацией функциональных возможностей организма и в том числе популяции тучных клеток. Поэтому обнаруженные нами изменения морфофункциональной активности тучноклеточной популяции ТК можно отнести к проявлениям срочной адаптации на воздействие гипотермии, которые характеризовались гипертрофией ТК, гранулолизисом и активной миграцией активированных юных форм ТК из кровяного русла. Миграцию ТК в легкие и их перераспределение при гипотермии можно рассматривать в рамках адаптационного синдрома. Аналогичные данные получили О.С. Арташян и соавт. (2012) на крысах, подвергнутых иммобилизационному стрессу. При воздействии стресса авторы наблюдали перераспределение и миграцию ТК в органы: число их увеличивалось в коже, желудке, кишечнике, печени, надпочечниках, в то время как в тимусе и костном мозге их количество значительно уменьшалось [9].
Заключение
Таким образом, активация ТК под влиянием гипотермии – закономерный процесс, так как ТК экспрессируют множество биологически активных веществ, которые стимулируют регенерацию легочной ткани после повреждения. ТК являются важными факторами, которые обеспечивают устойчивость дыхательной системы к воздействию гипотермии. Реакция ТК определяет ход процессов регенерации, изменений микроциркуляторного русла, восстановления микроокружения, питания паренхиматозных элементов и выработки адаптационных механизмов при действии гипотермии.
Библиографическая ссылка
Бобров И.П., Лепилов А.В., Гулдаева З.Н., Долгатов А.Ю., Алымова Е.Е., Крючкова Н.Г., Лушникова Е.Л., Молодых О.П. ТУЧНОКЛЕТОЧНАЯ ИНФИЛЬТРАЦИЯ ЛЕГКИХ КРЫС ПОСЛЕ ГИПОТЕРМИИ // Современные проблемы науки и образования. 2019. № 1. ;URL: https://science-education.ru/en/article/view?id=28446 (дата обращения: 01.08.2026).