


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
STRUCTURAL AND FUNCTIONAL DIVERSITY OF URBAN SOILS’ FUNGAL COMMUNITIES UNDER THE LOW TEMPERATURE CONDITIONS

Микробные сообщества, являясь одним из основных элементов почвенного покрова, играют ключевую роль в круговороте веществ в природе и являются важнейшим индикатором любых изменений в биосфере [1]. Большинство исследований свойств почвенной микобиоты в настоящее время проводится в условиях положительных температур почвы (с весны до осени) [2]. Однако в ряде работ показано, что и зимой отмечается активная жизнедеятельность почвенной биоты [3; 4].
До сих пор основное внимание в почвенно-микологических исследованиях уделялось изучению изменения видового разнообразия микобиоты, а исследований функционального разнообразия почвенных грибов практически не проводилось из-за отсутствия отработанных методов. Понятие функционального разнообразия определяется как общий совокупный результат взаимодействий организмов в экосистеме. Функциональное разнообразие организмов является показателем стабильности и устойчивости экосистемы [5]. Исследований изменения функционального разнообразия почвенной микобиоты при низких температурах до настоящего времени не проводилось.
Целью данной работы является оценка изменения структурного и функционального разнообразия микобиоты городских почв в условиях зимних температур. В фокусе данного исследования стоит вопрос изучения видовой и функциональной структуры сообществ микроскопических грибов городских почв при различных уровнях промораживания (-18, -2, +2 °C).
Объекты и методы
В качестве объекта исследования был выбран урбанозем (почвенно-мониторинговая площадка в СЗАО г. Москвы, ул. Виллиса Лациса). Содержание гумуса в верхнем горизонте А составляет 2,75%, рН (водн.)=6,4. Отбор и анализ почвенных образцов урбанозема проводился в начале ноября 2012 и 2013 гг. Лабораторные эксперименты проводили в термостате, где проводилось промораживание и оттаивание почвы в течение трех суток при температурах -18, -2, +2 °С. Температура -18 °С была взята как экстремальная; такое промораживание потенциально возможно для почв открытых пространств без снегового покрова. Температура -2 °С соответствует типичному значению для поверхностных горизонтов зональных дерново-подзолистых почв в зимний период. +2 °С – температура оттаивания почв исследуемого региона.
Оценку функционального разнообразия микроскопических грибов проводили методом мультисубстратного тестирования [6]. Набор субстратов, использованных в экспериментах, включал следующие вещества: арабиноза, галактоза, глюкоза, мальтоза, рибоза, сахароза, ксилоза, целлобиоза, аланин, цистеин, изолейцин, фенилаланин, гистидин, глутамин, лизин, серин, аспаргин, креатин, N-ацетилглюкозамин, дульцит, маннит, янтарная кислота. Приготовление планшетов с субстратами для анализов проводили самостоятельно. Предварительно в каждую лунку была внесена жидкая смесь среды Чапека (двойная стандартная концентрация, объем 100 мкл) с определённым органическим субстратом в концентрации 0,02 г/мл. Для проведения анализа из каждого смешанного образца готовили 25 мл почвенной суспензии с разведением 1:50, встряхивали на вортексе 2 мин, а затем центрифугировали в течение 3 мин (1000 оборотов/мин). Для предотвращения роста бактерий использовали раствор стрептомицина с концентрацией 70 мг/мл, 300 мкл которого смешивали с отобранным супернатантом. Приготовленную для анализа смесь вносили с помощью многоканального дозатора в лунки планшета с сухими субстратами в объёме 100 мкл. Для оценки роста грибов был использован кинетический подход: измерение оптической плотности (при 405 нм) вели каждые полчаса в течение 70-72 часов инкубации непосредственно в фотометре Sunrise (Tecan) при режиме встряхивания в течение 5 секунд перед измерением. Для сравнения полученных данных был использован метод трапеций для подсчета площадей под кривыми потребления субстратов. После регистрации в фотометре оптической плотности, проводили просмотр ячеек под оптическим микроскопом для оценки роста грибов и исключения возможности бактериального заражения. Из ячеек с наиболее интенсивным развитием грибов делали посев на агаризованную питательную среду Чапека (сахароза 30 г/л, натрия нитрат 2,0 г/л, калия гидрофосфат 1,0 г/л, магния сульфат 0,5 г/л, калия хлорид 0,5 г/л, железа сульфат 0,01 г/л, агар-агар 15,0 г/л) (с добавлением стрептомицина в качестве бактериального ингибитора).
Оценку видовой структуры сообществ культивируемых микроскопических грибов в лабораторных экспериментах с урбаноземом проводили методом посева из почвы на твердую агаризованную среду Чапека разведением 1:100. Все засеянные чашки инкубировали при температуре 25-27 °С в течение 7-10 дней, после чего проводили идентификацию культур. Определение видов производилось по культурально-морфологическим признакам [7]. Структуру, а также изменение функционального разнообразия грибных комплексов урбанозема характеризовали при помощи показателей обилия основных групп видов и индекса Шеннона.
Статистическая обработка данных проводилась с помощью программы Microsoft Office Excel 2007.
Результаты и обсуждение
В результате исследования установлено, что грибами исследуемого урбанозема потребляется довольно широкий спектр органических веществ: глюкоза, мальтоза, ксилоза, сахароза, целлобиоза, арабиноза, глутамин и аспарагин, маннит, N-ацетилглюкозамин. Промораживание почвы при -18 °С дает отчетливое снижение функционального разнообразия потребляемых субстратов. В то же время при обычной температуре промораживания почвы (-2 °С) существенных отличий в разнообразии потребляемых грибами субстратов не отмечается (табл. 1). Таким образом, серьезные изменения в функциональном разнообразии потребляемых субстратов потенциально могут произойти в зимний период в почвах открытых пространств, не защищенных снеговым покровом.
Таблица 1
Изменение индексов функционального разнообразия (Шеннона) грибных комплексов урбанозема в лабораторных экспериментах (70 часов инкубации)
|
Год |
Контроль |
-18 °С |
-2 °С |
+2 °С |
|
2012 |
3,34 |
1,98 |
- |
2,21 |
|
2013 |
3,68 |
- |
3,34 |
3,52 |
После промерзания исследуемых образцов почв при их оттаивании в лабораторных экспериментах (+2 °С) в целом наблюдается некоторое увеличение набора потребляемых субстратов по тенсивности развития на них грибов, при этом в наибольшей степени это видно по развитию грибов насилозе и N-ацетилглюкозамине (рис. 1).
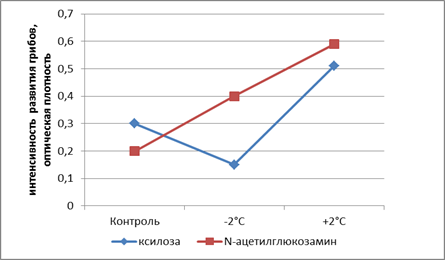
Это подтверждается и при подсчете площадей с помощью метода трапеций под кривыми потребления грибами субстратов. Данный метод применяется для оценки реакции популяций на воздействие экологических факторов, например для подсчета повременной суммарной численности клеток популяции в качестве интегрального способа оценки эффекта токсиканта [8]абл. 2, рис. 2).
Таблица 2
Значение площадей под кривыми потребления некоторых субстратов в лабораторных экспериментах (оптическая плотность)
|
Субстрат |
Исходный вариант |
При -2 °С |
При +2 °С |
|
Ксилоза |
5,0 |
3,4 |
9,8 |
|
N-ацетилглюкозамин |
3,3 |
6,3 |
13,6 |
Таким образом, показано, что в результате промораживания-оттаивания может происходить стимуляция развития грибов, потребляющих эти субстраты (рис. 2). Данный аспект интересен с точки зрения экологии городских почв, поскольку эти вещества содержатся в определенных компонентах экосистем. Источником поступления ксилозы в почву может быть растительный опад или вещество клеточных стенок ряда базидиальных грибов, поскольку ксилоза входит в состав полисахаридов гемицеллюлозы матрикса клеточной стенки. N-ацетилглюкозамин является мономером хитина – основного компонента экзоскелета членистоногих и ряда других беспозвоночных, а также входит в состав клеточной стенки грибов. Показано, что при -18 °С не происходит потребления N-ацетилглюкозамина, тогда как при -2 и +2 °С данный субстрат потребляется грибами активно. В экспериментах с урбаноземом 2013 г. отмечается даже увеличение интенсивности потребления N-ацетилглюкозамина с увеличением температуры (табл. 2).
А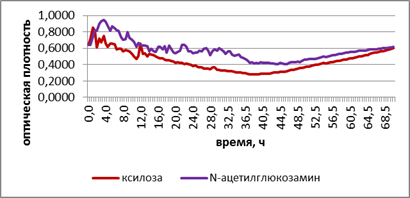
Б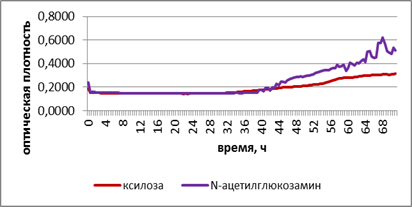
В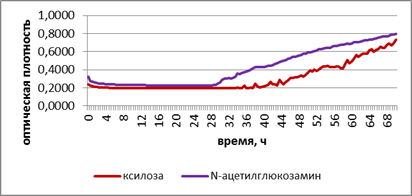
Рис. 2. Развитие грибов на ксилозе и N-ацетилглюкозамине при промораживании-оттаивании городских почв. А – потребление субстратов в исходном варианте. Б – потребление субстратов при -2 °С. В – потребление субстратов при +2 °С
Помимо прочего, одним из чувствительных показателей на температурное воздействие было изменение длительности лаг фазы при потреблении субстратов грибами. При промораживании-оттаивании в эксперименте наблюдалось отчетливое отличие по изменению времени начала потребления субстратов. По сравнению с промороженными почвами для оттаивающих наблюдалась меньшая лаг-фаза. Особенно четко это прослеживается по развитию грибов на аминокислотах (рис. 3).
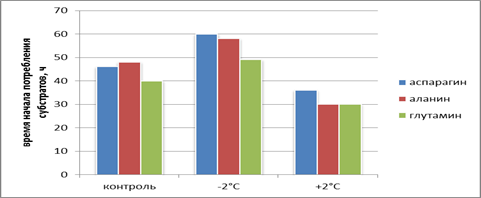
Рис. 3. Время начала потребления ряда органических субстратов грибами в экспериментах с промораживанием и оттаиванием урбаноземов
Таким образом, в результате оттаивания возможно более быстрое начало развития грибов, растущих на аминокислотах. Полностью объяснить наблюдаемые явления пока сложно. Возможно, более быстрое и активное потребление аминокислот определяется более выгодным использованием в стрессовых условиях более богатых по питательным веществам субстратов. Более быстрое начало потребления субстратов может быть связано со снятием явления фунгистазиса. Фунгистазиз определяется экзогенным (зависящим от внешних факторов) и конститутивным (внутренним) покоем грибных спор. В данной ситуации воздействия низких температур, возможно, могут работать оба механизма.
При анализе видовой структуры показано, что наибольшее ее изменение происходит в почвах при оттаивании (+2 °С), при этом выделяется ряд фито- и энтомопатогенных видов родов Beauveria, Fusarium (табл. 3).
Таблица 3
Видовой состав и обилие (%) культивируемых микроскопических грибов в урбаноземах в эксперименте замораживания-оттаивания (-2, +2 °С)
|
Виды |
Обилие, % |
||
|
Контроль |
-2 °С |
+2 °С |
|
|
Beauveria bassiana (Bals.-Criv.) Vuill. |
0,5 |
- |
3,1 |
|
Cladosporium sp. |
- |
- |
1,4 |
|
Clonostachys rosea (Link) Schroers, Samuels, Seifert & W. Gams |
- |
22,2 |
16,9 |
|
Fusarium graminearum (Schwein) Petch |
- |
4,9 |
6,5 |
|
Humicola sp. |
- |
14,6 |
- |
|
Mortierella sp. |
10,4 |
6,0 |
13,8 |
|
Mucor hiemalis Wehmer |
11,9 |
10,2 |
6,7 |
|
Penicillium citrinum Thom |
2,5 |
2,5 |
6,9 |
|
P. decumbens Thom |
- |
3,6 |
- |
|
P. janczewskii K.M. Zalessky |
1,8 |
14,4 |
- |
|
P. jantinellium Biourge |
- |
- |
16,8 |
|
P. melinii Thom |
47,4 |
- |
- |
|
P. simplicisimum (Oudem.) Thom |
11,6 |
- |
- |
|
Penicillium spp. |
0,5 |
- |
- |
|
P. waksmanii K.M. Zalessky |
- |
13,5 |
19,0 |
|
Phoma sp. |
0,6 |
1,0 |
- |
|
Trichoderma polysporum (Link) Rifai |
4,2 |
- |
- |
|
Verticillium tenerum Nees |
2,8 |
- |
- |
|
Стерильный мицелий |
5,8 |
7,1 |
8,9 |
При выделении на питательную среду в контрольном варианте без температурного воздействия доминируют виды рода Penicillium (P. melinii, P. simplicisimum и др.), а также представители р. Mortierella. Показано, что они сохраняются в почвах и после замораживания-оттаивания, но виды рода Penicillium выделяются в меньшей степени. В экспериментах отмечалось, что после промораживания из почв обильно (до 22,2%) начинает выделяться Clonostachys rosea. Из почв после промораживания также выделяются Humicola sp., P. janczewskii, Fusarium graminearum. При оттаивании после замораживания, из образцов урбаноземов обильно выделяется Clonostachys rosea (16,9%), а также P. jantinellium, P. waksmanii (табл. 3).
Таким образом, в модельных экспериментах выявлено, что процессы промораживания-оттаивания могут оказывать влияние на функциональное разнообразие комплексов микроскопических грибов, что выражается в снижении индексов разнообразия потребляемых субстратов, задержки их потребления грибами в результате промораживания образцов. По сравнению с промороженными почвами при оттаивании отмечалась меньшая лаг-фаза начала потребления ряда субстратов почвенными грибами. Особенно четко это прослеживается по развитию грибов на аминокислотах и сильнее проявляется после промораживания при низкой температуре -18 °С. В оттаивающих почвах отмечается активное развитие грибов на таких субстратах, как ксилоза и N-ацетилглюкозамин. Стоит также отметить, что одновременно как при выделении на наборе субстратов (на планшетах), так и при выделении посевом на питательную среду в урбаноземах при оттаивании отмечалось некоторое увеличение выделения фитопатогенных грибов Fusarium graminearum и энтомопатогенных грибов Beauveria bassiana.
Заключение
В ходе проведенного исследования показано, что снижение функционального разнообразия наблюдается при сильном промораживании (-18 °С), а при температурах, характерных для зональных зимних условий (-2 °С), отличий не выявлено. При оттаивании (+2 °С) промороженных почв происходит быстрое начало потребления грибами ряда аминокислот и интенсивности нарастания мицелия. В отличие от летнего периода, когда в урбаноземах доминировали представители р. Penicillium, зимой часто выделялись представители р. Clonostachys. После оттаивания исследованных городских почв отмечено более частое выделение из них энтомо- и фитопатогенных грибов (Beauveria bassiana, Fusarium graminearum).
Библиографическая ссылка
Тепеева А.Н., Марфенина О.Е. СТРУКТУРНОЕ И ФУНКЦИОНАЛЬНОЕ РАЗНООБРАЗИЕ МИКОБИОТЫ ГОРОДСКИХ ПОЧВ В УСЛОВИЯХ ПОНИЖЕННЫХ ТЕМПЕРАТУР // Современные проблемы науки и образования. 2017. № 5. ;URL: https://science-education.ru/en/article/view?id=26842 (дата обращения: 29.06.2026).