


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
FEATURES OF THE FORMATION OF THE DOMINANT SPECIES OF ABOVE-GROUND MASS IN THE STEPPIES FACIES OF THE MINUSINSK BASIN

Южно-Минусинская котловина относится к числу наиболее освоенных районов юга Красноярского края. Природные условия определяются резко континентальным климатом и расположением в центральной части Азиатского материка вдали от морских бассейнов. Настоящие ковыльные степи Минусинской котловины, по классификации Е.М. Лавренко [5], входят в состав так называемых островных степей Сибири, а флора представлена большим числом высокогорных, горностепных и монгольских элементов. Левобережная часть Енисея в пределах юга Минусинской котловины известна как Койбальская степь, она является уникальным типом степей. Растительный покров Койбальской степи составляют крупнодерновинные злаки (ковыльные и овсецовые), которые выступают в качестве коренных сообществ. Мелкодерновинные степи в данном регионе разнообразны и занимают большие площади. К ним можно отнести также холмисто-увалистые настоящие четырехзлаковые степи, выделенные В.В. Ревердатто [8]. Основу их составляют злаки: типчак валисский – Festuca valesiaca, тонконог гребенчатый – Koeleria cristata,, мятлик кистевидный – Poa botryoides, и змеевка растопыренная – Cleistogenes squarrosa. Почвенный покров исследуемой территории представлен среднемощными малогумусными карбонатными черноземами в комплексе со степными солонцами [7].
Климат котловины довольно теплый: средняя температура июля составляет 19 °С, среднегодовая положительна 0,9 °С, выпадает 350-450 мм осадков в год, максимальное количество приходится на летние месяцы (50-60%). Наибольшее количество осадков выпадает в июле и августе (за исключением сухих лет). Весна и начало лета обычно засушливы – все это накладывает отпечаток на рост и развитие растений. Зимние осадки составляют 20-30%. Высота снежного покрова не превышает 19 см, снег часто сдувается в лога и лощины, что ужесточает перезимовку степных растений.
Объекты и методы исследования
Для оценки биологической продуктивности используются данные общего запаса растительного вещества и его состава отдельных видов. Основные методы определения биологической продуктивности, которых придерживаемся, изложены в работах [6; 9]. Надземная масса растений учитывалась на площадках в 0,25 м2 методом укосов в 3-5-кратной повторности. Проводилось подробное описание площадок, на каждой из них определялось проективное покрытие каждого вида и сделаны зарисовки в горизонтальной проекции. Сбор образцов проводился в период максимального развития травостоя (конец июля, начало августа). Зеленую массу разбирали по видам. Все пробы растительного вещества высушивались в течение 8-12 часов при температуре 105 °С до абсолютно сухого веса, затем взвешивались. Ошибка средней для данных надземной массы составила ±10-16%.
Исследования проводились на полигон-трансекте Новониколаевского стационара, который пересекает несколько пространственно сопряженных фаций с мелкогрядовыми и плоскоравнинными формами рельефа (рис. 1).
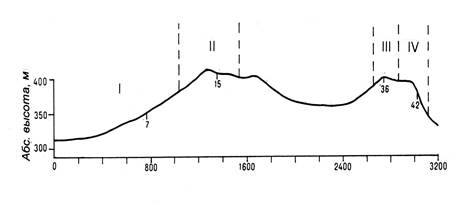
Рис. 1. Основные фации полигона-трансекта Койбальской степи
Фации: I – мелкодерновинно-злаково-ковыльная с караганой южного склона; II – петрофитно-разнотравно-ковыльно-типчаковая выровненной поверхности; III – осоково-вострецово-ковыльная с караганой вершинной поверхности; IV – разнотравно-осоково-овсецово-ковыльная денудационного склона северной экспозиции [3].
Изменение экологических условий на полигон-трансекте отражается в многообразии растительных сообществ, характеризующихся видовым составом и продуктивностью. Резкие контрасты в пространственном распределении растительного покрова создаются микрорельефом, представленным гребневидными вершинами, уступами на южных склонах и нивальными нишами на северных склонах. Расположение фаций отражает изменение структуры растительного покрова территории и зависит от характера освещения, теплообеспеченности, увлажнения, засоления, петрофитности – т.е. факторов, которые, в свою очередь, на ограниченной территории являются функцией пространственной вариации рельефа. В пределах полигон-трансекта со сменой геоморфологических поверхностей (склоновой денудации, транзита и аккумуляции) растительный покров представлен рядом последовательно сменяющих друг друга сообществ, охватывающих несколько фаций.
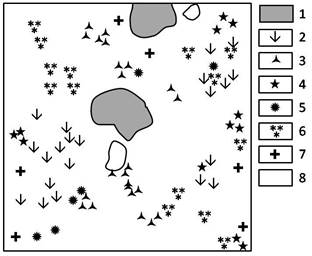
Рис. 2. Горизонтальная проекция мелкодерновинно-злаково-ковыльного с караганой сообщества южного склона, 2010 г. (ф. I):
1 - Stipa Krylovii, 2 - Aneurolepidium pseudoagropyrum, 3 - Caragana pygmaea, 4 - Carex pediformis,
5 - Cleistogenes squarrosa, 6 - Artemisia frigida,
7 - Artemisia commutate, 8 - Helictotrihom desertorum
Фация I – мелкодерновинно-злаково-ковыльная с караганой южного склона широко распространена и занимает пологие склоны. Проективное покрытие сообщества в разные годы имеет значительные флуктуации по метеоусловиям года. Для условий 2010 г. проективное покрытие 60-70%. Строение травяного полога 3-ярусное. Изучаемую фацию можно отнести к сообществам, достаточно заполненным видами, при этом насыщенность зависит от размера площади, на 1 м2 насчитывается до 18 видов. На площади 0,25 м2 в мелкодерновинно-злаково-ковыльном сообществе зафиксировано 8 видов растений (рис. 2). Господствующим видом крупнодерновинных злаков для этой фации является ковыль Крылова – Stipa Krylovii. Одновременно доминирующими видами из мелкодерновинных злаков являются: типчак – Festuca valesiaca, змеевка растопыренная – Cleistogenes squarrosa, тонконог гребенчатый – Koeleria cristata, мятлик оттянутый – Poa botryoides, вострец ложнопырейный – Aneurolepidium pseudoagropyrum, овсец пустынный – Helictotrihom desertorum. Фактически на фации I численность дерновинных злаков не снижается, но проявляются периодические флуктуации в разные годы. Доминантами из разнотравья являются полынь холодная – Artemisia frigida, подмаренник настоящий – Galium verum, скабиоза Фишера – Centaurea fisheri, василистник малый – Thalíctrum mínus, эдельвейс эдельвейсовидный – Leontopodium leontopodioides, из бобовых видов – карагана карликовая – Caragana pygmaea.
Фации II и III занимают вершинные поверхности. Следует отметить, что в этих фациях вегетация растений обычно начинается рано (середина апреля) и продолжается до самой поздней осени (середина октября), а некоторые дерновинные злаки и осоки уходят под зиму. У типчака, ковылей и осоки перезимовавшие листья после схода снега продолжают существовать и функционировать до появления новых побегов, а потом постепенно отмирают. Таким образом, к концу мая – началу июня появляются побеги новой генерации [2]. Экологические условия петрофитно-разнотравно-ковыльно-типчаковой выровненной поверхности (ф. II) на вершине полигон-трансекта наименее благоприятны для жизни растений (каменистые почвы). На гривах травостой наиболее разрежен и слагается в основном типчаком и петрофитным разнотравьем. Проективное покрытие 50-60 в отдельные годы до 80%. На 1 м2 насчитывается 21 вид. На основной поверхности фации ковыль вместе с овсецом пустынным образуют фрагментарный ярус высотой от 40-50 до 65 см. На площадках 0,25 м2 насчитывается до 8 видов растений.
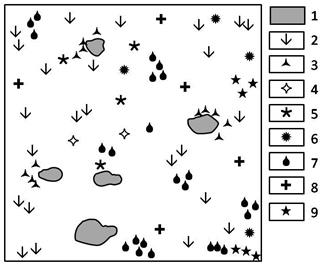
Рис. 3. Горизонтальная проекция осоково-вострецово-ковыльного с караганой сообщества вершинной поверхности, 2010 г. (ф. III):
1 - Stipa Krylovii, 2 - Aneurolepidium pseudoagropyrum, 3 - Caragana pygmaea, 4 - Oxytropis grandiflora,
5 - Euphorbia borealis, 6 - Cleistogenes squarrosa,
7 - Festuca valesiaca, 8 - Artemisia commutate,
9 - Carex pediformis
Фация III – осоково-вострецово-ковыльная с караганой вершинной поверхности, не отличается особой сложностью. Травостой высотой 50-60 см представлен ковылем Крылова – Stipa Krylovii. На фации пятнами выделяются осочково-вострецовые сообщества с большим участием полыни замещенной и холодной – Artemisia commutate и frigida, вострецом ложнопырейным – Aneurolepidium pseudoagropyrum, здесь же присутствуют мелкодерновинные виды: типчак – Festuca valesiaca и тонконог гребенчатый – Koeleria cristata. Кроме преобладающих на этой поверхности южных черноземов, пятнами встречаются солонцеватые почвы, на которых произрастает осока твердоватая – Carex duriuscula, из разнотравья – подмаренник настоящий – Galium verum, из бобовых – карагана карликовая – Caragana pygmaea. Проективное покрытие в 2010 г., на осоково-вострецово-ковыльной фации достигает 70%. Строение травяного полога 3-ярусное. Первый ярус составляет 50-70, второй – 40-50 и третий – 10-30 см. На 1 м2 насчитывается 24 вида. На горизонтальной поверхности фации III (0,25 м2) насчитывается 9 видов растений (рис. 3), количество видов может увеличиваться до 12 в зависимости от условий тепла и влаги.
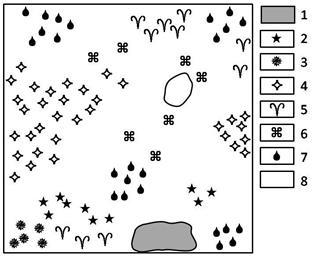
Рис. 4. Горизонтальная проекция разнотравно-осоково-овсецово-ковыльного сообщества склона северной экспозиции, 2010 г. (ф. IV).
1 - Stipa Krylovii, 2 - Carex pediformis, 3 - Aster alpinus, 4 - Oxytropis grandiflora, 5 - Poa botryoides,
6 - Bupleúrum auréum, 7 - Festuca valesiaca,
8 - Helictotrihom desertorum
Фация IV – разнотравно-осоково-овсецово-ковыльная денудационного склона северной экспозиции. Отличительными свойствами фации IV являются мезофильный характер этой фации, а также самое большое по трансекту видовое разнообразие растений. Горизонтальное сложение травостоя довольно равномерное (рис. 4). На общем фоне пятнами выделяются осоково-типчаковые сообщества с преобладающим участием типчака – Festuca valesiaca. В сообществах северного склона на щебнистых почвах произрастает овсец пустынный – Helictotrihom desertorum. В благоприятные годы высота травостоя достигает 60-80 см. Этот травяной полог состоит из целого ряда мобильных видов: крестовник цельнолистный – Senecio integrifolius, колокольчик сибирский – Campánula sibírica, осока стоповидная – Carex pediformis, ирис русский – Iris ruthenica, астра альпийская – Aster alpinus, вероника седая – Veronica incana. Сложившийся состав травостоя разнотравно-осоково-овсецово-ковыльного сообщества продолжает долго вегетировать, т.е. зеленые вегетативные части растений можно обнаружить здесь до середины октября. Это растения, корневая система которых простирается глубже 10-15 см – Caragana pygmaea, Oxytropis grandiflora, Artemisia frigida.
Результаты исследования
Стационарные исследования позволяют получить материалы по распределению общих запасов зеленой массы и доминирующих видов растений в фациях полигон-трансекта, а также выявить пространственно-временные формирования видов в изучаемых сообществах.
Изменение запасов растительных сообществ на профиле в разные годы обусловлено различными проявлениями жизнедеятельности: одни растения – со стабильным доминированием, другие испытывают флуктуации, третьи уходят из фитоценоза при сильном выпасе или пожаре, а четвертые усиливают свои позиции с увеличением солонцеватости [4]. Из состава степной флоры для анализа были взяты растения, играющие наиболее выраженную роль в сложении травостоя – доминанты, содоминанты и остальные виды, входящие в состав общей зеленой массы травостоя, которые показаны в таблице.
Долевое участие, продуктивность доминирующих видов и запасы общей зеленой массы в фациях полигон-трансекта Койбальской степи
|
Виды растений |
Фации |
|||||||
|
I |
II |
III |
IV |
|||||
|
% |
г/м2 |
% |
г/м2 |
% |
г/м2 |
% |
г/м2 |
|
|
Ковыль Крылова – Stipa Krylovii |
21 |
47 |
29 |
84 |
29 |
76 |
14 |
30 |
|
Овсец пустынный – Helictotrihom desertorum |
21 |
48 |
15 |
41 |
10 |
27 |
14 |
30 |
|
Типчак – Festuca valesiaca |
6 |
8 |
3 |
9 |
5 |
14 |
6 |
13 |
|
Тонконог гребенчатый – Koeleria cristata |
1 |
3 |
1 |
4 |
8 |
21 |
5 |
10 |
|
Змеевка растопыренная - Cleistogenes squarrosa |
1 |
2 |
3 |
9 |
12 |
32 |
2 |
4 |
|
Мятлик оттянутый – Poa botryoides |
1 |
3 |
12 |
34 |
2 |
6 |
3 |
6 |
|
Вострец - Aneurolepidium pseudoagropyrum |
5 |
11 |
2 |
7 |
7 |
9 |
11 |
25 |
|
Осока стоповидная – Carex pediformis |
9 |
20 |
4 |
12 |
5 |
14 |
19 |
42 |
|
Карагана карликовая – Caragana pygmaea |
12 |
26 |
8 |
23 |
7 |
18 |
4 |
8 |
|
Остальные виды |
25 |
55 |
21 |
58 |
13 |
33 |
23 |
50 |
|
Общая зеленая масса |
223 |
281 |
260 |
218 |
||||
Существующая в пространстве неоднородность в распределении надземной массы зависит от структуры различных растительных сообществ и эколого-биологических особенностей слагающих видов. Для фаций I, II, III характерно заметное участие кустарников, составляющих 7-12% надземной массы. Общей особенностью структуры видового состава всех фаций является доминирование нескольких видов. Среди них только два вида всегда представительны в надземной массе: из крупнодерновинных злаков доминант – ковыль Крылова – Stipa Krylovii (14-29%) и субдоминант – овсец пустынный – Helictotrihom desertorum (10-21%), из мелкодерновинных злаков типчак – Festuca valesiaca (3-9%) и змеевка растопыренная – Cleistogenes squarrosa (1-12%), для других видов характерна приуроченность к определенным местообитаниям. В мезофитных условиях разнотравно-осоково-овсецово-ковыльного денудационного склона северной экспозиции в верхней части (ф. IV) происходит возрастание в надземной массе осоки стоповидной – Carex pediformis (19%), востреца ложнопырейного – Aneurolepidium pseudoagropyrum (11%). Участие полыни холодной – Artemisia frigida играет заметную роль только в фациях, приуроченных к щебнистым местообитаниям (ф. II), и на ее долю приходится до 17% [1].
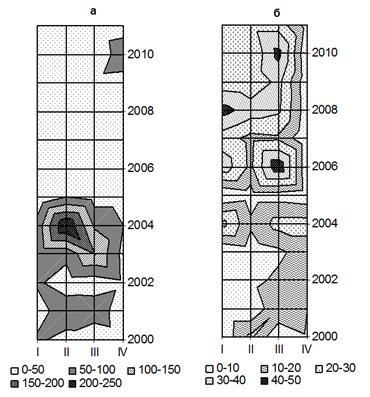
Рис. 5. Пространственно-временные модели (2000-2011 гг.) крупнодерновинных и мелкодерновинных видов (г/м2) на фациях полигон-трансекта: а – ковыль Крылова (Stipa Krylovii), б – типчак валисский (Festuca valesiaca)
Построенные графические модели в пространственно-временном ряду, где каждая фация имеет вполне определенную многолетнюю динамику доминирующих видов (рис. 5а). Прослеживается четкая ритмичность, в первую очередь обусловленная своеобразием гидротермических условий конкретной фации. Каждая фация достигает максимального накопления фитомассы видов в конкретный, только для них срок. Состояние травостоя полностью зависит от количества осадков, температуры и влияния пожаров. Из изучаемых лет наиболее благоприятными для развития ковыля Крылова были 2003-2004 гг., о чем свидетельствуют максимальные величины запасов - от 200 до 250 г/м2 на фациях I и II. Количество осадков за 2 месяца активной вегетации (июль, август) составило в 2004, 2005 и 2006 гг. 122,6, 87,6 и 129,5 мм соответственно. Таким образом, наиболее благоприятный температурный режим при сочетании тепла и влаги в 2003-2004 гг., по-видимому, и объясняет значительное увеличение запасов ковыля. В 2005 г. наступает резкий спад запасов ковыля до 50 г/м2 в связи засушливым летом и малым количеством осадков - 87,6 мм (июль и август). Несмотря на большое количество осадков в 2006 г. (129,5 мм в июле и августе), запасы ковыля Крылова достигали всего 50 г/м2, возможно, повлияли весенние пожары. Запасы ковыля в благоприятные 2003-2004 гг. на фации I и II в 5 раз выше, чем запасы типчака, это связано не только с термическими условиями, но и с физиологическими свойствами развития самих растений. Поведение запасов типчака прямо противоположное, по сравнению с видом ковыля Крылова (рис. 5б). Самое высокое по годам участие и запасы типчака до 50 г/м2 наблюдаются в 2006 и 2010 гг. на фации III - осоково-вострецово-ковыльной с караганой и в 2008 г. мелкодерновинно-злаково-ковыльной с караганой ф. I, с благоприятным сочетанием тепла и влаги. В засушливый 2005 г. запасы типчака минимальны (от 1 до 10 г/м2) на всех исследуемых фациях, и это сочетается с очень низким жизненным состоянием из-за недостатка влаги в почве и высокой температуры воздуха.
Заключение
Полученные результаты по продуктивности степей Минусинской котловины позволяют выявить тенденции развития степных сообществ в многолетнем режиме функционирования. Система центральноазиатских типов степей подтверждается в основном содержанием видов растительности, по создаваемой продуктивности, численности и проективному покрытию сообществ биологической продуктивности полигон-трансекта.
Показана роль злаков растительных сообществ как критерий отношения сообществ к типу настоящих степей. Значительная роль принадлежит крупнодерновинным злакам: Helictotrihom desertorum, Stipa Krylovii, и они занимают лидирующее положение. При смене режимных условий мелкодерновинные злаки: Festuca valesiaca, Koeleria cristata - также остаются в числе доминирующих.
Прослеживается четкая ритмичность доминирующих видов растительности, обусловленная своеобразием гидротермических условий в пространственно-временном ряду конкретной фации.
Библиографическая ссылка
Дубынина С.С. ОСОБЕННОСТИ ФОРМИРОВАНИЯ ДОМИНИРУЮЩИХ ВИДОВ НАДЗЕМНОЙ МАССЫ В СТЕПНЫХ ФАЦИЯХ МИНУСИНСКОЙ КОТЛОВИНЫ // Современные проблемы науки и образования. 2016. № 3. ;URL: https://science-education.ru/en/article/view?id=24796 (дата обращения: 02.07.2026).