


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
ANNUAL DYNAMICS OF WATER EXCHANGE AND PHOTOSYNTHETIC PIGMENTS OF CONIFEROUS SPECIES OF SPRUCE IN LOW LIGHT LEVEL CONDITIONS

Основным показателем водного статуса растений является транспирационная способность ассимиляционных органов лесных видов. Интенсивность транспирации в лесных экосистемах по величине и физиологической значимости имеет первостепенное значение, особенно в условиях низкого уровня освещенности, как это имеет место под пологом древостоя сомкнутого ельника. Многофункциональной роли световой энергии в физиологии древесных растений посвящено большое число исследований, основоположников в этой области: [1, 5, 6]. По данным [6], освещенность под пологом сомкнутого древостоя ельника на склонах южной экспозиции соответствует 10–16 %; на северных – 6–8 %; на восточных и западных склонах – 9–11 % света от солнечной энергии на открытых местах. При этом возобновление подроста и его состояние вполне удовлетворительное. В связи с этим возникает необходимость изучения динамики расхода воды в процессе транспирации и других показателей водного обмена: ОСВ, ОПВ, оводненности хвои лесных растений, по-видимому, имеющих специфичность адаптивности разных видов хвойных к недостатку света. Наряду с количественной оценкой поступающей ФАР и УФ радиации, важна роль температуры и влажности воздуха, часто являющие факторами, регулирующими фотосинтетические процессы и состояние водного обмена растений.
Целью данной работы является характеристика годовой динамики показателей водного обмена и динамики фотосинтетических пигментов хвои на фоне регистрируемых параметров микроклимата в вегетационных опытах с длительным притенением саженцев видов рода Picea.
Объекты и методы
Вегетационные опыты являются наиболее рациональной моделью для изучения ответных адаптивных реакций при действии экстремальных факторов среды, одним из которых является низкий уровень освещенности ювенильных хвойных пород. Объекты исследований – пятилетние саженцы местных видов ели аянской (Piceaajanensis (Lind. etGold.) Fisch. exCarr.), ели корейской (PiceakoraiensisNakai.) и интродуцента – ели обыкновенной (Piceaabies (L.) Karst.). Исследования проводили в однотипных почвенных и климатических условиях вегетационных опытов с 2009 по 2011 год. При постановке опыта саженцы видов ели переносили из лесного питомника, с хорошо инсолируемого склона юго-западной экспозиции в вегетационный домик под пленочным покрытием. Использовали притенение саженцев пологом для снижения освещенности до 30 % в 2009 году, 16,5 % в 2010 г и 8 % – в 2011 году. В контрольном варианте освещенность саженцев регистрировалась под полиэтиленовой пленкой вегетационного домика. Интенсивность транспирации измеряли методом быстрого взвешивания [4]. Относительное содержание воды (ОСВ) определяли на изолированной хвое, помещенной в дистиллированную воду в закрытых чашках Петри до полного насыщения тканей хвои в течение 24 часов и рассчитывали по методике [9]. Относительную потерю воды (ОПВ) из хвои саженцев определяли по [10]. Количество пигментов в хвое саженцев ели определяли по методике [8]. Показатели микроклимата Фар и УФ – радиацию, температуру и влажность воздуха регистрировали 5, 15 и 25 числа каждого месяца, с использованием прибора ТКМ – ПКМ – 42».
Результаты и обсуждение
Характеристика показателей водного статуса саженцев ели
В годовой динамике интенсивности транспирации саженцев ели наблюдается тенденция снижения этого показателя с 2009 года по 2011 год (рис. 1), за исключением летнего периода 2010 года, аномально жаркого по климатическим параметрам. В июле 2010 года показатели транспирации хвои были наиболее высокими, а в 2011 году транспирация хвои всех видов ели была значительно ниже, чем предыдущие 2009 и 2010 годов.
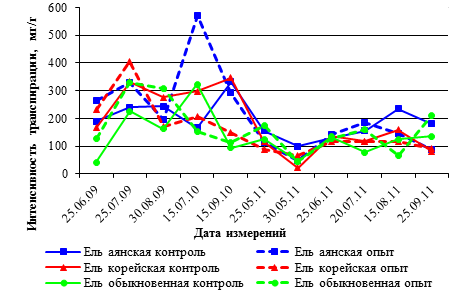
Рис. 1. Интенсивность транспирации хвои видов ели (мг/г сырого веса в час) в вегетационных опытах 2009–2011гг.
В июне 2009 года, через 15 дней после пересадки саженцев из питомника и их притенения, интенсивность транспирации у всех видов ели в условиях низкой освещенности больше, чем в контроле у саженцев ели аянской на 73 мг/г; ели корейской на 64 мг/г; обыкновенной – на 85 мг/г. Такие различия между опытом и контролем по интенсивности транспирации хвои сохраняются в течение всего летнего периода. К концу августа месяца 2009 года интенсивность транспирации у саженцев местных видов – ели аянской и корейской в контроле и в опыте имеют близкие значения, в отличие от интродуцента ели обыкновенной, у которой интенсивность транспирации на 147 мг/г в опыте больше, чем в контроле (табл. 1).
Таблица 1
Интенсивность транспирации саженцев ели по дням наблюдений, 2009 г.
|
Растения |
Вариант опыта |
Дата наблюдений |
||||
|
25.06.09 |
25.07.09 |
Разница* |
30.08.09 |
Разница |
||
|
15 дней |
45 дней |
65 дней |
||||
|
Ель |
Контроль |
190 |
243 |
244 |
||
|
Опыт |
263 |
330 |
87 |
197 |
47 |
|
|
Ель |
Контроль |
166 |
332 |
276 |
||
|
Опыт |
230 |
405 |
73 |
172 |
104 |
|
|
Ель обыкновенная |
Контроль |
41 |
216 |
162 |
||
|
Опыт |
126 |
330 |
114 |
309 |
147 |
|
* – разница в опыте по отношению к контролю.
В 2011 году так же, как и в 2010 году, интенсивность транспирации хвои саженцев всех видов ели в опыте и контроле имеет близкие значения, но в условиях длительного притенения и значительного снижения освещенности в вегетационном опыте 2011 года уровень интенсивности транспирации значительно ниже, чем в 2009 году, особенно у местных видов ели аянской и корейской к середине сентября 2011 года (табл. 2).
Таблица 2
Интенсивность транспирации саженцев ели по дням наблюдений, 2011 г.
|
Растения |
Вариант опыта |
Дата наблюдений |
||||
|
25.05.11 |
25.06.11 |
20.07.11 |
15.08.2011 |
25.09.2011 |
||
|
15 дней |
45 дней |
70 дней |
95 дней |
120 дней |
||
|
Ель |
Контроль |
156 |
128 |
155 |
235 |
180 |
|
Опыт |
114 |
141 |
184 |
145 |
88 |
|
|
Ель |
Контроль |
116 |
135 |
118 |
159 |
181 |
|
Опыт |
88 |
115 |
116 |
115 |
92 |
|
|
Ель обыкновенная |
Контроль |
124 |
134 |
77 |
125 |
136 |
|
Опыт |
174 |
126 |
160 |
67 |
211 |
|
Годовая динамика показателей относительного содержания воды в тканях хвои саженцев ели также имеет тенденцию к снижению ОСВ хвои у всех изучаемых пород к 2011 году.
Уже в третьей декаде мая 2011 года ОСВ саженцев всех видов ели меньше в варианте опыта с низким уровнем освещенности, по сравнению с контролем и с показателями ОСВ в 2009–2010 годах. У разных видов ели уровень ОСВ хвои различается незначительно. Величина значений ОСВ у всех видов ели варьирует в разные годы исследований от 61 до 97 %. Самыми низкими показатели ОСВ были в хвое саженцев ели обыкновенной (61,9–78.0 %) в течение 2011 года (рис. 2).
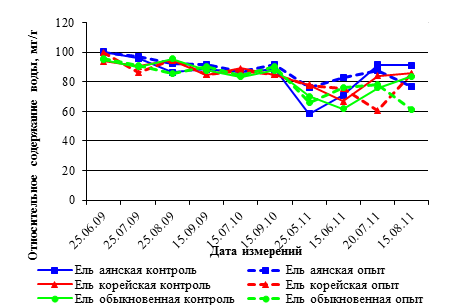
Рис. 2. Годовая динамика ОСВ,% в хвое саженцев видов р. Picea в условиях низкого уровня освещенности, 2009–2011 гг.
В ходе длительной адаптации (в нашем случае три вегетационных сезона) саженцев ели к условиям пониженного освещения разница ОСВ в хвое в контроле и в опыте незначительна. По средним значениям показатели ОСВ соответствовали в 2009 г. – 91,8 %; в 2010 году – 86.6 %; в 2011 г. – 75,3 % (табл. 3).
Годовая динамика относительной потери воды в хвое саженцев ели также имеет тенденцию снижения показателей ОПВ с 2009 года к 2011 году.
В летний период 2010 года самые высокие показатели ОПВ – 2,54 мг/г в контроле и 3,42 мг/г в опыте были отмечены у саженцев ели аянской в середине августа месяца. В течение трех лет исследований значения ОПВ хвои, как правило, больше у саженцев всех видов ели в опытном варианте, чем в контроле. Процент увеличения ОПВ хвои в опыте по отношению к контролю в 2010 году в июле был самым высоким и соответствовал от 30 % в хвое ели аянской до 68 % в хвое ели обыкновенной.
Таблица 3
Относительное содержание воды в хвое саженцев ели по дням наблюдений, (%)
|
Растения |
Вариант опыта |
Дата наблюдений |
||
|
25.07.09 |
15.07.10 |
25.05.11 |
||
|
45 дней |
45 дней |
45 дней |
||
|
Ель |
Контроль |
96 |
86,4 |
85,5 |
|
Опыт |
97 |
87,2 |
76,2 |
|
|
Ель |
Контроль |
90,6 |
89,2 |
77,9 |
|
Опыт |
86 |
87,9 |
77,2 |
|
|
Ель обыкновенная |
Контроль |
90,3 |
83,5 |
70,2 |
|
Опыт |
91,3 |
84,2 |
65,9 |
|
Скорость потери воды из хвои у всех видов саженцев ели в 2011 году была в 5–10 раз меньше, чем в 2009 году, и в 12–17 раз меньше, чем в 2010 году (рис. 3).
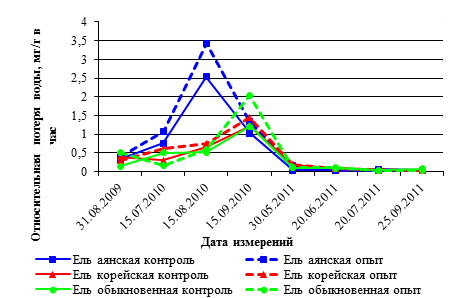
Рис. 3. Годовая динамика ОПВ в хвое видов р. Picea в вегетационных опытах 2009–2011 гг. (в мг/г сырого веса в час)
В летний период 2011 года процент увеличения ОПВ по отношению к контролю был значительно ниже, чем в предыдущие годы, и соответствует 9 % в хвое ели аянской, 28 % в хвои ели корейской и 30 % в хвое ели обыкновенной. Саженцы разных видов ели по показателям ОПВ хвои располагаются по убывающей последовательности: ель аянская – ель обыкновенная – ель корейская (табл.4).
В вегетационных опытах в разные годы влажность почвы варьировала от 15 до 45 %. Влажность почвы в 2010 году была значительно меньше, чем в предыдущие годы. Значения влажности почвы, как правило, больше в опыте, чем в контроле.
В вегетационных опытах 2009 и 2010 гг. влажность хвои саженцев местных видов ели в вегетационных опытах 2009 г. и 2010 г. соответствует 54,6 – 68 ,2 %, за исключением ели обыкновенной, у которой влажность хвои в июне месяце была несколько больше – 73,7 %.
Таблица 4
Относительная потеря воды хвои саженцев ели по дням наблюдений,
мг/г сырого веса в час
|
Растения |
Вариант опыта |
Дата наблюдений |
|||
|
15.08.10 |
15.09.10 |
20.06.11 |
20.07.11 |
||
|
75 дней |
80 дней* |
40 дней |
70 дней |
||
|
Ель |
Контроль |
2,54 |
1,03 |
0,033 |
0,061 |
|
Опыт |
3,42 |
1,34 |
0,038 |
0,038 |
|
|
Ель |
Контроль |
0,65 |
1,22 |
0,085 |
0,034 |
|
Опыт |
0,67 |
1,45 |
0,088 |
0,047 |
|
|
Ель обыкновенная |
Контроль |
0,52 |
1,21 |
0,086 |
0,03 |
|
Опыт |
0,59 |
2,03 |
0,12 |
0,043 |
|
Таким образом, анализ транспирационной способности хвои саженцев ели в течение трех вегетационных сезонов показал, что резкая смена освещенности открытого пространства в питомнике на условия притенения в опыте вызывает снижение интенсивности транспирации хвои в опыте, по сравнению с контролем. Длительное затенение саженцев всех видов ели в опыте сопровождается снижением интенсивности транспирации хвои с 405 мг/г в час в 2009 году до 141 мг/г в час в 2011 году (по максимальным значениям). Хвойные растения имеют высокую адаптивную способность к очень низкому уровню освещенности и длительному его воздействию.
Годовая динамика фотосинтетических пигментов в хвое саженцев ели
В июле 2009 года у всех изучаемых видов ели количество пигментов по их сумме в варианте опыта с притенением в 1,3–1,4 раза меньше, чем в контрольном варианте (табл. 5). Процент снижения пигментов (Аа+в+кар.) по их сумме в опыте по отношению к контролю в хвое саженцев ели аянской равен 11,8 %, в хвое ели корейской – 22,9 % и в хвое ели обыкновенной – 24,9 %. Сумма хлорофиллов (Аа+в) также меньше в опыте, чем в контроле. Процент снижения суммы хлорофиллов в хвое саженцев ели аянской равен15,6 %, в хвое ели корейской – 25,3 % и в хвое ели обыкновенной – 27,9 %.
В июле 2010 года и в июле 2011 года количество пигментов в хвое всех видов ели в опытном варианте, наоборот, увеличивается по отношению к их содержанию в контрольном варианте. В 2010 году процент увеличения пигментов в опыте по отношению к контролю соответствует 31,5 % (табл. 5). Процент увеличения пигментов в опыте по отношению к контролю в 2011 году в хвое ели корейской находится в пределах 27–35 %. В хвое ели обыкновенной процент увеличения количества пигментов в опыте по отношению к контролю был еще больше – 65–70 % (табл. 5).
Таблица 5
Количество пигментов в хвое видов рода Picea в условиях длительного затенения
|
Вариант |
∑ Аа+в |
∑ Аа+в+кар |
||||
|
Контроль |
Притенение |
% снижения (-), увеличения (+) |
Контроль |
Притенение |
% снижения (-), увеличения (+) |
|
|
25.07.2009 г. |
||||||
|
Ель аянская |
1,92 |
1,62 |
(-)15,6 |
2,20 |
1,94 |
(-)11,8 |
|
Ель корейская |
1,98 |
1,48 |
(-)25,6 |
2,31 |
1,78 |
(-)22,9 |
|
Ель обыкновенная |
1,36 |
0,98 |
(-)27,9 |
1,65 |
1,24 |
(-)24,9 |
|
15.07.2010 г. |
||||||
|
Ель корейская |
1,21±0,13 |
1,74±0,08 |
(+)30,5 |
1,44±0,15 |
2,03±0,09 |
(+)29,1 |
|
Ель обыкновенная |
1,35±0,09 |
1,97±0,14 |
(+) 31,5 |
1,61±0,11 |
2,28±0,17 |
(+) 29,4 |
|
25.07.2011 г. |
||||||
|
Ель корейская |
1,85±0,12 |
2,85±0,13 |
(+)35,1 |
2,19±0,16 |
3,00±0,20 |
(+)27,0 |
|
Ель обыкновенная |
0,71±0,05 |
2,39±0,10 |
(+) 70,3 |
0,87±0,06 |
2,60±0,06 |
(+)65,5 |
Установлено, что наиболее сильные изменения в фотосинтетическом метаболизме углерода и синтезе пигментов осуществляются в первое время после смены освещенности открытого питомника на вариант с притенением в первый летний период вегетации 2009 года, при этом происходит снижение уровня пигментов по их сумме в хвое ели аянской на 11,8 %, у ели корейской на 22,9 %, а в хвое ели обыкновенной – на 24,9 % по отношению к контролю. Это означает, что регуляторные процессы ФСМУ и, как следствие – синтез первичных продуктов фотосинтеза [3] и пигментов, начинается сразу с момента снижения освещенности и уменьшения количества продуктов фотохимических реакций, протекающих в хлоропластах, что в некоторой степени подтверждается исследованиями, проведенными на листьях картофеля в условиях резкой смены освещенности на притенение растений [7].
В ходе длительной адаптации (в нашем случае во второй и третий вегетационные сезоны 2010 г. и 2011 г.) к условиям очень низкого уровня освещенности происходит увеличение синтеза пигментов в хвое ели корейской и ели обыкновенной на 29,1 и 29,4 %. В условиях длительного пониженного освещения, вероятно, происходит уменьшение синтеза транспортной формы углеводов хвойных – сахарозы [3], и растения изыскивают в процессе адаптации альтернативные пути сохранения экспортной функции листа.
Следует отметить, что приспособление хвойных древесных видов к недостатку полного спектра световой энергии может сопровождаться как анатомо-морфологическими, так и фотохимическими и биохимическими преобразованиями, поэтому так трудно выявить механизм и длительность периода фотоадаптаций. В течение проведенных нами трехлетних вегетационных опытов на одних и тех же саженцах видов ели, с постепенно снижающимся уровнем освещенности опытных растений, нами установлено, что наиболее сильным воздействием на саженцы хвойных была резкая смена условий освещенности на притенение (до 30 % от освещенности в контроле). В начале первого этапа (2009 г.) эксперимента происходит снижение интенсивности транспирации, снижение уровня пигментов, как по их сумме (А а+в+ кар.), так и по сумме хлорофиллов (А а+в). В 2010 году у саженцев ели аянской в опыте отмечена высокая скорость потери воды (ОПВ) хвои, а также снижение на 10 % относительного содержания воды в хвое (ОСВ). В вегетационном сезоне 2011 г. уровень освещенности в опытном варианте был идентичен освещенности под пологом сомкнутого ельника. Влияние длительного притенения сопровождалось снижением интенсивности транспирации хвои (см. табл. 2), снижением показателей ОСВ в среднем с 90 % до 70 % значительным снижением показателей ОПВ и оводненности хвои всех видов саженцев ели по отношению к контрольным данным и показателям водного обмена саженцев ели в 2009 и 2010 годах
В условиях длительной адаптации (в течение трех вегетационных летних сезонов) к условиям очень низкого освещения саженцы хвойных изыскивают альтернативные пути сохранения фотосинтетической функции хвои. Одной из таких проявлений адаптивной способности следует считать значительное увеличение в летний период 2010 и 2011 годов фонда фотосинтетических пигментов в хвое саженцев ели в опыте по отношению к контролю.
Выводы
Для хвойных растений нами впервые установлено, что в хвое саженцев видов ели существует регуляторный комплекс реакций фотосинтетического метаболизма с регуляцией показателей водного статуса хвои и реагирующий на изменение интенсивности солнечной радиации. Ключевыми моментами, вероятно, являются регуляторные механизмы водного обмена и фотосинтетические процессы в хлоропластах, снижение или увеличение синтеза пигментов. В результате ФСМУ растения реагируют на притенение при резкой смене световых условий на теневые – снижением уровня пигментов, а при длительном затенении – адаптивные реакции поддерживаются повышением уровня пигментов в хвое и регуляцией водного обмена.
В отличие от покрытосемянных растений, у хвойных видов рода Picea ответные адаптивные реакции на длительное затенение проявились наиболее сильно (по изменению показателей водного статуса и уровню пигментов) только на третий год вегетации саженцев в условиях очень низкой инсоляции.
Библиографическая ссылка
Козина Л.В., Титова М.С., Репин Е.Н. ГОДОВАЯ ДИНАМИКА ПОКАЗАТЕЛЕЙ ВОДНОГО ОБМЕНА И ФОТОСИНТЕТИЧЕСКИХ ПИГМЕНТОВ ХВОЙНЫХ ВИДОВ ЕЛИ В УСЛОВИЯХ НИЗКОГО УРОВНЯ ОСВЕЩЕННОСТИ // Современные проблемы науки и образования. 2016. № 3. ;URL: https://science-education.ru/en/article/view?id=24765 (дата обращения: 02.07.2026).