


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
INTERACTION AMONG RED WOOD ANTS AND PASSERINE BIRDS

Комплексные взаимодействия между таксономически далекими группами видов исследуются чаще всего в пределах одной гильдии. Например, была продемонстрирована трофическая конкуренция между между зерноядными пустынными грызунами и муравьями [3]. Полевые исследования взаимодействия между членами разных гильдий до сих пор редки. Так, не только не проводилось систематических исследований, но даже не высказывалось предположений о характере межвидовых взаимодействий таких существенных компонентов лесных экосистем, представляющих различные крупные таксономические группировки, как рыжие лесные муравьи и воробьинообразные птицы.
Рыжие лесные муравьи группы Formica rufa принадлежат к “видам-инженерам”, преобразующим среду обитания для многих видов животных и растений. В лесных биоценозах ассоциации муравейников могут занимать сотни гектаров, а биомасса достигать 10 кг на 1 га [2]. До сих пор исследовалось взаимодействие рыжих лесных муравьев, главным образом, с беспозвоночными [9]. Лишь единичные исследования посвящены анализу межвидовых отношений рыжих лесных муравьев с позвоночными животными. Недавно исследованы комплексные взаимодействия этих насекомых с мелкими млекопитающими и выяснено, что они связаны многоплановыми отношениями, включающими трофические взаимодействия, элементы топической конкуренции и синойкию. Важно отметить, что муравьи, выступая как фактор беспокойства, существенно снижают численность и роющую активность мышевидных грызунов на своей территории, а также препятствуют их миграционной активности [1]. Выявлены поведенческие адаптации мышевидных грызунов, позволяющие им эффективно охотиться на агрессивных и опасных насекомых [5]. Изучены отдельные аспекты взаимодействия рыжих лесных муравьев с воробьинообразными птицами. В природных экспериментах при предоставлении синицам выбора между подкормочными площадками, содержащими муравьев и аналогичными площадками без муравьев, синицы не только чаще посещают изолированные от муравьев площадки, но и проводят там больше времени [6]. С помощью учетов в искусственных гнездах и соответствующих наблюдений было показано, что беспокойство и атаки со стороны рыжих лесных муравьев заставляют птиц (несколько видов синиц и мухоловок) выбирать для мест гнездования деревья, где этих насекомых нет [7]. Присутствие муравьёв приводит к снижению жизненно важных показателей (вес, упитанность) у птенцов пищухи Certhia familiaris [4]. В то же время известно, что воробьинообразные птицы могут включать в свой рацион жалящих перепончатокрылых, а большие синицы используют их для выкармливания птенцов.
Данная работа является первым шагом в комплексном исследовании экологических и этологических аспектов взаимодействия рыжих лесных муравьев и воробьинообразных птиц. В ней поставлены следующие задачи:
1. исследовать население воробьинообразных птиц кустарникового яруса на территории, контролируемой муравьями, и прилегающем контрольном участке; 2. сравнить охотничье поведение разных видов воробьиных птиц по отношению к рыжим лесным муравьям; 3. исследовать влияние присутствия муравьев на сроки выведения птенцов у мухоловки-пеструшки;
Материалы и методы
Исследования проводились в мае - июле 2012-2014 гг. на территории Приобской лесостепной провинции в 30 км от г. Новосибирска в березовом лесу с березово-осиновыми, осиново-березовыми участками, посадками ели, сосны и лиственницы, с обильным подлеском черемухи. Было выбрано крупное поселение муравьев Formica aquilonia, занимающее площадь более 30 га и насчитывающего более 500 гнезд. Выделен экспериментальный участок площадью 0,7 га, на котором динамическая плотность муравьев составляла 60 ± 29 экз./10 дм2/мин. Контрольные участки площадью 0,5 и 0,7 га располагались за пределами муравьиного поселения. Экспериментальный и контрольный участок выбирались так, чтобы древесно-кустарниковая растительность была сходной, и основным фактором, отличающим эти участки друг от друга, было присутствие крупного поселения рыжих лесных муравьев.
Исследование населения птиц кустарникового яруса проводилось при помощи стандартного метода отловов паутинными сетями на экспериментальном и первом контрольном участке. Сети закреплялись на двух вертикальных шестах высотой 2,5 м, что позволяло отлавливать птиц, обитающих или кормящихся в кустарниковом ярусе. Сети устанавливались в 6.00 и снимались в промежуток с 11 до 13 часов (в зависимости от погодных условий) и проверялись 1 раз в 20-30 минут. Всего проведено 30 отловов в июне 2013 и в июне 2014 гг. По результатам отловов рассчитывался коэффицент общности Жаккара для видовых составов на экспериментальном и контрольном участках по формуле K=с/(a+b-с), где а и b - количество видов, обнаруженных на экспериментальном и первом контрольном участках соответственно, с - количество общих видов). Такое количество данных позволяет оценить качественные различия в видовом составе птиц на экспериментальном и контрольном участках.
Для изучения сроков заселения и выведения птенцов на экспериментальном участке было развешено 15 искусственных гнездовий для мелких птиц с круглым летком диаметром 32 мм, на втором контрольном участке— 9. Птицы заселяли искусственные гнездовья в конце мая. Каждое гнездовье проверялось один раз за сезон (14 июня 2012 г., 21 июня 2013 г. и 20 июня 2014 г.) после начала выклева на обоих участках, фиксировалась стадия развития выводка: кладка или птенцы. Соотношение стадий развития на контрольном и экспериментальном участках сравнивалось при помощи критерия Манна-Уитни. Данный метод позволяет сравнить сроки заселения искусственных гнездовий и требует только однократного учёта всех развешенных гнездовий, в отличие от альтернативных методов (требующих наблюдения дуплянок в течение всего гнездового сезона). Ранее, характеризуются ранним выклевом птенцов. Соотношение кладок и выводков птенцов на участке характеризует среднюю дату откладки яиц.
Для исследования охотничьего поведения разных видов птиц использовали прозрачный пластиковый контейнер (300х300х500 мм), содержащий две присады в виде наклонных жердочек, закрепленных концами на дне и стене арены. В контейнер последовательно помещали 50 муравьев и птицу, отловленную с помощью паутинной сети. Эксперименты проводили в затемненной комнате с искусственным освещением. Поведение птицы фиксировалось видеокамерой в течение 10 минут. После однократного тестирования птицы возвращались на участок отлова. При обработке видеоматериала поминутно подсчитывали количество «успешных» (заканчивающихся попаданием в муравья клювом) и «неудачных» (заканчивающихся промахом или незавершенных) атак. Для каждой птицы рассчитывали общее количество атак за 10 минут и успешность (отношение суммы “успешных” атак к общему количеству, умноженная на 100%). Всего протестировано 18 птиц 8 видов (4 особи мухоловки-пеструшки Ficedula hypoleuca, по 3 особи чечевицы Carpodacus erythrinus и большого пестрого дятла Dendrocopos major, по 2 особи садовой камышовки Acrocephalus dumetorum, рябинника Turdus pilaris и белобровика T. iliacus и по одной особи большой синицы Parus major и поползня Sitta europaea).
Результаты
На экспериментальном участке отловлена 61 птица 17 видов, на контрольных участках отловлено 42 птицы, относящихся к 12 видам (таблица). На территории, контролируемой муравьями, отловлено большее количество видов, по сравнению с территорией, свободной от муравьев (17 и 12). На контрольном участке не были отмечены D. major (большой пестрый дятел), Erithacus rubecula (зарянка), Luscinia luscinia (соловей обыкновенный), Phylloscopus collybita (пеночка-теньковка), P. montanus (буроголовая гаичка). Видовой состав птиц на опытном и контрольных участках сходен, индекс общности Жаккара равен 0,63. Как на экспериментальном, так и на опытном участках встречаются характерные для данного биотопа виды, такие как F. hypoleuca (мухоловка-пеструшка), S. minula (славка-завирушка), T. pilaris (рябинник), Ph. phoenicurus (горихвостка-лысушка).
Оказалось, что наиболее активно и успешно охотились поползень (86 % успешных атак), большая синица (66%) и белобровик (91%). Мухоловка-пеструшка и чечевица проявляли меньший интерес к муравьям, попытки атаковать их редко заканчивались успехом (0 - 38% успешных атак у разных особей мухоловки, 0 - 25% для разных особей чечевицы).

Рис. 1 Количество и процент успешных атак у разных видов птиц. Большой пестрый дятел обозначен как б.п. дятел, мухоловка-пеструшка — как мухоловка.
Развешанные искусственные гнездовья на экспериментальном и контрольном участке заселялись исключительно мухоловкой-пеструшкой. В 2012 году было заселено 8 и 5, в 2013 – 14 и 8, в 2014 - 10 и 6 из 15 гнездовий на экспериментальном и 9-и на контрольном участке соответственно. За все время учетов на экспериментальном участке было учтено достоверно больше (критерий Манна-Уитни, Uemp=0, p<0.05) выводков на поздней стадии развития по сравнению с контрольным участком (рис.2)
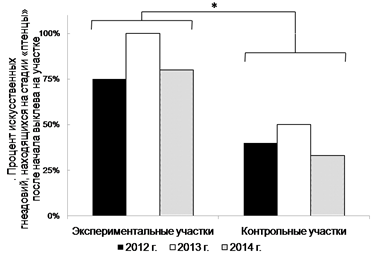
Рис. 2. Процент искусственных гнездовий, находящихся на стадии «птенцы» после начала выклева на участке. * p<0.05, критерий Манна-Уитни.
Обсуждение результатов
По данным отловов сообщество мелких воробьиных в выбранном биотопе включает 18 видов птиц, при этом на территории, контролируемой муравьями, отловлено больше видов, чем на участке, свободном от них. На данном этапе исследования мы не можем связать это с влиянием рыжих лесных муравьев вследствие микромозаичности сообщества птиц и недостаточного количества наших данных. По всей видимости, для выявления тонких различий в населении птиц необходимо применять методы учета гнездящихся пар. Применение подобных методов может выявить неявные различия, обусловленные влиянием рыжих лесных муравьев, например различия в плотности населения, кормовом поведении или гнездовом успехе. Интересно отметить, что сходные результаты были получены при исследовании взаимодействия муравьев и мелких млекопитающих: состав сообществ сходен на территориях, контролируемых и не контролируемых муравьями, а проявляемые эффекты касаются непосредственного взаимодействия животных [1, 5].
Птицы всех 8 выбранных для тестирования видов продемонстрировали в охотничье поведение по отношению к муравьям. Для каждого вида выборка невелика (от 1 до 4 особей), и полученные данные следует рассматривать как предварительные, но и на данном этапе различия в суммарном количестве и успешности атак птиц на муравьев можно объяснить экологической специализацией. Так, высокий процент успешных атак у поползня можно объяснить тем, что ползающие по стенкам арены муравьи позволяют ему использовать характерный для вида способ питания – склевывание со стенок. Подобные преимущества имеют и дрозды (рябинник и белобровик), тогда как зерноядные птицы (чечевица) не могут успешно охотиться в условиях нашего эксперимента. Промежуточное положение в этих условиях занимают насекомоядные птицы - садовая камышовка и мухоловка-пеструшка. Мухоловки-пеструшки демонстрируют большой индивидуальный разброс в успешности атак (от 0 до 78%). Следует отметить, что рыжие лесные муравьи входят в природные рационы многих видов птиц, а в некоторых случаях могут доминировать в питании взрослых особей, что было показано для мухоловки-пеструшки [10]. Межвидовые отношения птиц и муравьев могут иметь различный характер в зависимости от природной зоны и конкретных взаимодействующих видов. В умеренной зоне продемонстрированы поведенческие адаптации больших синиц к взаимодействию с рыжими лесными муравьями [6]. Как оказалось, в выбранном модельном сообществе взаимное влияние муравьев и птиц перспективно исследовать на примере мухоловки-пеструшки, так как только этот вид заселил искусственные гнездовья, не оставив ни одного свободного. На экспериментальном участке птенцы вылупляются достоверно раньше, чем на контрольных. Это дает возможность предположить, что активность муравьев в кронах деревьев оказывает влияние на процесс гнездования и выведения птенцов. У мухоловки-пеструшки от момента откладывания яиц до выклева птенцов проходит 12-13 дней, от момента выклева до вылета 13-18 дней. Мы предполагаем, что разница в сроках выклева птенцов мухоловки-пеструшки на опытном и контрольном участках обусловлена разным сроком заселения искусственных гнездовий. Стоит при этом отметить, что рыжие лесные муравьи выбирают для своих поселений хорошо прогреваемые, освещенные и продуктивные участки леса [2], и не исключено, что воробьинообразные птицы в первую очередь стремятся занять те же территории, вследствие сходства экологических предпочтений с рыжими лесными муравьями. Возможно, что мухоловки-пеструшки заселяют прежде всего занятые муравьями территории, и птенцы у таких пар появляются раньше. Вследствие своей массовости в начале мая, сами муравьи могут быть привлекательным пищевым объектом для только что прилетевших птиц. В это время муравьи в огромном количестве появляются на поверхности муравейников, прогревая их, и доступны для склевывания. При этом известно, что диета мухоловок-пеструшек может состоять из муравьев на 90% [8, 10]. Вопрос о том, наблюдается ли у них ухудшение жизненных показателей птенцов, по аналогии с результатами, полученными на пищухах [4], и если да, то компенсируется ли это более ранними сроками развития, требует дальнейшего исследования.
Заключение
Видовой состав воробьиных птиц в кустарниковом и травянисто-кустарниковом ярусах на территории, контролируемой муравьями, включает виды, характерные для исследуемых местообитаний. На участке, занятом крупным поселением муравьев, отмечено 17 видов, на контрольных участках – 12, при этом видовой состав сообществ сходен. Охотничье поведение по отношению к муравьям на экспериментальных аренах демонстрировали в той или иной мере все 8 исследованных видов птиц. Наиболее успешно атаковали муравьев большая синица, поползень и дрозд – белобровик. Искусственные гнездовья как на экспериментальном, так и на контрольном участках, занимает исключительно мухоловка-пеструшка. На экспериментальном участке птенцы вылупляются достоверно раньше. Можно полагать, что активность муравьев в кронах деревьев оказывает влияние на процесс гнездования и выведения птенцов.
Исследования поддержаны Российским научным фондом (грант № 14-14-00603).
Рецензенты:
Резникова Ж.И., д.б.н., проф., зав. лабораторией поведенческой экологии сообществ института систематики и экологии животных, г. Новосибирск;
Юдкин В.А., д.б.н., старший научный сотрудник лаборатории зоомониторинга, Институт систематики и экологии животных Сибирского отделения Российской академии наук, г. Новосибирск.
Библиографическая ссылка
Маслов А.А., Пантелеева С.Н. ВЗАИМОДЕЙСТВИЕ РЫЖИХ ЛЕСНЫХ МУРАВЬЕВ И ВОРОБЬИНЫХ ПТИЦ // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/en/article/view?id=23839 (дата обращения: 04.07.2026).