


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE DISPERSION OF THE ELECTRICAL CONDUCTIVITY OF THE RAT BRAIN DURING HYPOTHERMIA AND THE INTRODUCTION OF UREA

Одним из признаков, характеризующих гипотермические состояния, является уровень физиологической активности клеток различных тканей. Чем ниже температура, при которой сохраняется физиологическая активность, тем меньше риск летального исхода при прочих равных условиях. Поэтому поиск режимов, при которых критическая температура полного подавления физиологической активности снижена по сравнению с нормой, является одним из путей решения проблемы создания безопасных гипотермических состояний. Нервная ткань весьма чувствительна к различным воздействиям и одним из компонентов ее реакции является изменение электрического сопротивления [2]. Известно [4], что при гипотермии (26 °С) происходит значительное увеличение сопротивления тканей мозга (4,7% в ретикулярной формации, 7% в гиппокампе).
Раннее нами было показано, что мочевина оказывает защитное действие при глубокой гипотермии: при введении мочевины перед охлаждением животного температура тела, при которой электроэнцефалограмма становится изоэлектрической, снижается [1]. Мочевина постоянно присутствует в организме, но не усваивается ЦНС и в физиологических концентрациях не токсична для мозговой ткани. Синтезируется этот осмотик в различных органах, в том числе и в мозге, являясь одним из конечных продуктов обмена млекопитающих. Связываясь с белками, мочевина может оказывать влияние на их функциональную активность и создавать осмотическое давление, влияя на водно-солевой баланс в клетках мозга. Одной из возможных причин влияния мочевины на электрическую активность мозга является ее способность вызывать перераспределение воды и ионов между нейронами и экстраклеточным пространством за счет осмотических эффектов. Эти эффекты могут влиять на электропроводность ткани мозга.
Целью работы явилось исследование частотной зависимости активной и реактивной компонент импеданса мозга крыс при гипотермии на фоне введения мочевины. Для этого фиксировали изменения пассивных электрических свойств мозговой ткани: регистрация активных и реактивных составляющих импеданса мостовым методом (трехэлектродный метод) и регистрация изменения потенциала с помощью вольтметра (четырехэлектродный метод).
Методика исследования
Животные. Опыты проведены на 12 крысах-самцах линии Вистар весом 180-200 г, содержащихся на обычном рационе в условиях вивария.
Наркоз. Все опыты проведены на животных наркотизированных тиопенталом в дозе 40 мг на кг веса тела.
Гипотермия. Снижение температуры тела (34, 25, 20, 19,5°С) осуществляли с помощью полиэтиленовых мешков, наполненных битым льдом. Наркотизированное животное помещали на мешки со льдом и периодически измеряли температуру тела ректально ртутным термометром. Время охлаждение составляло около 1,5 часа, а время согревания до нормотермии около 1 часа.
Введение мочевины. Мочевину вводили внутрибрюшинно в дозе 3мМ на 100 г веса тела.
Измерение электрических характеристик мозга мостовым методом. Измерительная система состоит из моста переменного тока Р5021, нуль-индикатора Ф582 и генератора переменных импульсов ГЗ-111, в качестве вольтметра - измеритель нелинейных искажений С6-7 - активная (Rw) и реактивная (Хw) составляющая импеданса. Суть измерения состоит в том, что необходимо уравновесить мост, регулируя ёмкость и сопротивления в измерительном плече моста, а в другое плечо включается исследуемый объект [6].
Измерение импеданса четырёхэлектродным методом. Наиболее радикальным способом устранения погрешности, связанной с поляризацией электродов, является разделение электродов на токовые и измерительные, то есть четырёхэлектродный метод. При этом, первые используются только для пропускания тока через ткань, а вторые - для отведения возникающих в ткани потенциалов, пропорциональны её специфическому импедансу. Относительные величины (процентные изменения) функциональных сдвигов мозга, измеренных мостовым методом, должны оказаться заниженными, т.к. они отнесены к суммарному сопротивлению ткани и сопротивлению электродов [2]. Особенно сильно возрастает электродная ёмкость и сопротивление на низких частотах, и неучёт этого обстоятельства может дать ложную частотную зависимость для тканевого сопротивления, как это иногда имеет место и делает результаты сомнительными [7]. Полученные данные анализировали в программе «STATISTICA».
Результаты исследования и обсуждение.
На Рис.1 и 2 приведены графики частотной зависимости Rw и Хw при гипотермии на фоне внутрибрюшинного введения мочевины. Само по себе введение мочевины слабо повлияло на Rw. Охлаждение организма при введении мочевины сначала приводит к некоторому увеличению Rw: при температуре тела 30 и 25°С Rw заметно выше, чем до начала охлаждения. При температуре тела 19,5°С кривая дисперсии Rw практически совпадает с начальной кривой при tr = 34°С. Таким образом, в отличии от контроля гипотермия слабо влияет на активную компоненту и уменьшает реактивную на 15% на частоте 1000 Гц при 25°С по сравнению с 34°С. Введение мочевины привело к температурной зависимости Rw мозга, активная компонента несколько увеличивается, а реактивная уменьшается. При этом зависимость реактивной компоненты от температуры тела при введении мочевины существенно сильнее при 25°С на частоте 1000 Гц реактивная компонента уменьшается на 42% по сравнению с 34°С.
Введение мочевины приводит к небольшому изменению Хw. При снижении температуры тела происходит дальнейшее уменьшение Хw. Следует отметить, что эти изменения более существенны, чем в контрольных опытах. Углубление гипотермии до 19,5°С, также как и в случае Rw, приводит к приближению величины реактивной компоненты к начальной кривой. Однако этот эффект в случае Rw меньше чем Хw.
Отсюда можно сделать вывод, что: а) введение мочевины оказывает влияние на импеданс мозга (как на Rw так и Hw); б) введение мочевины также оказывает влияние на динамику импеданса при гипотермии.
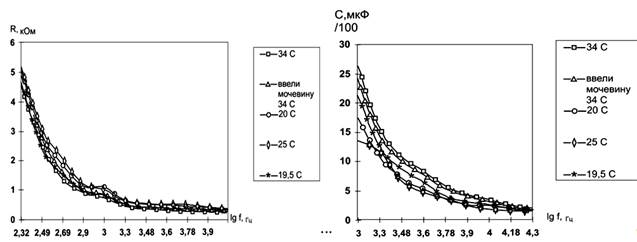
Рис.1.(а, б). Графики зависимости активной составляющей импеданса от частоты после введения мочевины в динамике гипотермии.
Изменения Rw указывает, видимо, на изменение омического сопротивления мозга при введении мочевины. Одной из причин такого влияния может быть изменение соотношения объемов экстраклеточного и внутриклеточного пространства. Увеличение омического сопротивления объясняется уменьшением объема экстраклеточного пространства. С другой стороны, поскольку зависит также и от емкости, увеличение Rw можно было бы объяснить уменьшением емкости.
Введение мочевины, во всяком случае, на первых этапах, могло бы привести к уменьшению экстраклеточного пространства за счет осмотического эффекта, а это должно привести к уменьшению омического сопротивления, что и наблюдается в наших опытах. Это уменьшение объема экстраклеточного пространства не связано с набуханием мозга, которое может наблюдаться при глубокой гипотермии вследствие подавления работы ионных насосов.
На рис. 2 (а, б и в) приведены частотные зависимости потенциалов регистрирующих электродов (V1 и V2) при гипотермии на фоне введения мочевины. Видно, что введение мочевины приводит к снижению потенциала в точках V2 и V3 в области 1000- 10000 Гц.
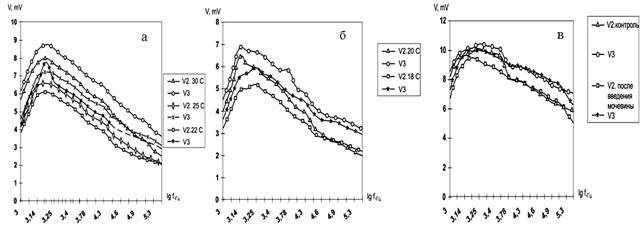
Рис.2. Графики зависимости потенциала в точках V2 и V3 от частоты после введения мочевины в динамике гипотермии.
Снижение температуры тела приводит к дальнейшему снижению этих потенциалов. Следует отметить, что разность потенциалов, подаваемая на токовые электроды, не зависит от частоты и, следовательно, изменение потенциала измеряющего электрода возможно только в том случае, если происходит изменение профиля потенциала в образце.
Исходя из схемы расположения электродов в мозге (рис.3.) и соотношения потенциалов в точках 2, 3 и 4 можно на качественном уровне предложить следующую схему распределения потенциалов в образце: поскольку электроды 1 и 4 токовые и, следовательно, заполяризованы, вблизи этих электродов происходит существенное падение потенциала из-за большого сопротивления заполяризованных электродов. Можно предположить, что сопротивление R1 и R2 значительно превосходит сопротивление R и, таким образом, ток, текущий между электродами 1 и 4 определяется суммой сопротивлений R1 и R2.
Рис.3. Схема распределения потенциалов в образце, где 1 - сопротивление между электродами 1 и 2. R2 - сопротивление между электродами 3 и 4. R = сопротивление ткани мозга, расположенной между регистрирующими электродами.
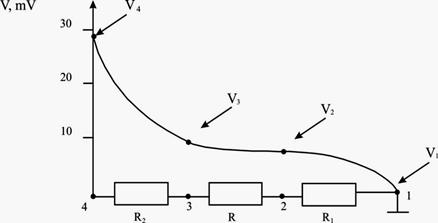
Как уже отмечалось выше, разность потенциалов V4 – V1 в условиях опыта постоянна и не зависит ни от частоты переменного тока, ни от температуры тела. Следовательно, разность потенциалов V3 - V2 в этих условиях равна:
V3 -V2 = I R, (1)
где I - ток текущий в цепи.
Предполагается, что
![]() , (2)
, (2)
При изменении температуры мозга изменяется сопротивление R. Однако, поскольку мы предполагаем, что R намного меньше (R1 + R2), то изменение R слабо влияет на величину I. Поэтому V3 — V2 оказывается пропорционально R. Таким образом, увеличение V3 - V2 соответствует увеличению R. Снижение температуры тела приводит к увеличению V3 - V2 , что свидетельствует об увеличении сопротивления (R) ткани мозга. Гипотермия слабо влияет на активную компоненту и уменьшает на 15% реактивную (на частоте 1000 Гц при 25°С по сравнению с 34°С). При введении мочевины активная компонента несколько увеличивается, а реактивная снижается. При этом зависимость реактивной компоненты от температуры тела при введении мочевины существенно сильнее (при 25°С на частоте 1000 Гц реактивная компонента уменьшается на 42% по сравнению с 34°С), то есть увеличение согласуются с результатами, полученными с использованием мостового метода [4].
Выводы
На основании анализа полученных нами данных можно сделать следующие выводы: а) в биологических системах наблюдается существенная частотная зависимость как активной так и реактивной составляющей импеданса; б) введение мочевины, как и ожидалось, привело к некоторому осмотическому эффекту: увеличивается омическое сопротивление за счет изменения отношения экстраклеточного и внутриклеточного пространства. Также наблюдается связанное с этим изменения и в электрических характеристиках импеданса; в) при измерениях четырехэлектродным методом также наблюдается частотная и температурная зависимость потенциала. При снижении температуры происходит снижении величины потенциала в точках V2 и V3. Что также согласуется с данными полученными при измерениях мостовым методом.
Библиографическая ссылка
Пиняскина Е.В., Абдурахманов Р.Г. ДИСПЕРСИЯ ЭЛЕКТРОПРОВОДНОСТИ ТКАНИ МОЗГА КРЫС ПРИ ГИПОТЕРМИИ НА ФОНЕ ВВЕДЕНИЯ МОЧЕВИНЫ // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/en/article/view?id=23771 (дата обращения: 04.07.2026).