


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
MORPHOFUNCTIONAL STATE OF THE PINEAL GLAND OF DIFFERENT SPECIES OF FOREST VOLE WITH THE SHORTENING OF THE DAYTIME AND HIGH DENSITY OF THE POPULATION

У лесной полёвки проявляется различная скорость полового созревания [9]. У вида красно-серая полёвка участие в размножении сеголеток позднелетней генерации определяется продолжительностью светлого времени суток, тогда как у красной полёвки – плотностью популяции. Высокая её плотность вызывает стресс. В реализации фотозависимых и опосредованных стрессом механизмов реализации программы онтогенеза принимает участие шишковидная железа [7]. Сдвиг морфометрических показателей активности пинеальной секреции выявлен у двухмесячных неполовозрелых самцов красно-серой полёвки позднелетней генерации [2]. При моделировании стресса на высокую плотность популяции красно-серой полёвки также обнаружились морфологические признаки гиперфункции шишковидной железы [5]. Установлено, что у крыс кариометрические показатели активности интерстициальных эндокриноцитов яичек изменялись синхронно с пинеалоцитами шишковидной железы, гипоталамическими нейроэндокринными клетками супрахиазматических и паравентрикулярных ядер, регулирующими выработку гонадолиберина и опосредующими через ретино-гипоталамическую проекцию влияние света и стрессоров на шишковидную железу, гонадотропными эндокриноцитами гипофиза, чувствительными к гонадолиберину и пинеальному антигонадотропному гормону [4]. В качестве морфологических маркёров оценки активности шишковидной железы, кроме гистохимической реакции на мелатонин и кариометрических показателей пинеалоцитов, используют их органеллометрические показатели, изменения ультраструктурной организации ядер, ядрышек, эндоплазматического ретикулума, комплекса Гольджи, лизосом, митохондрий, эндокринных везикул, синаптических лент, сфер, везикул с конкрециями [1, 3, 6, 8]. Каково функциональное состояние шишковидной железы, оказывающей влияние на выбор реализуемой программы онтогенеза у сеголеток разных видов лесной полёвки весенне-летней и позднелетней генераций при высокой плотности популяции, до настоящего времени остаётся не изученным.
Цель работы – оценить морфофункциональное состояние шишковидной железы у сеголеток-самцов красной полёвки и красно-серой в зависимости от продолжительности светлого времени суток и плотности популяции.
Материал и методы исследования
Работа выполнена на двухмесячных сеголетках весенне-летней и позднелетней генераций в соответствии с «Правилами проведения работ с использованием экспериментальных животных» (1977). Детёныши-самцы, рождённые от изъятых из природной среды самок, содержались при естественном освещении и выводились из эксперимента в 1000–1130 ч путём декапитации. После отсепаровывания кожи, перерезки в области лобного шва, симметричного кругового вскрытия в направлении от лобной части к затылочной, отламывания костей свода черепа шишковидная железа отделялась от мозговых оболочек. На 2 ч выделенный материал погружался в смесь 4% параформальдегида и 2,5% глютаральдегида на 0,1М какодилатном буфере (рН 7,4, 4⁰С), в течение 2 ч постфиксировался в 1% OsO4 на 0,1М какодилатном буфере (рН 7,4, 4⁰С), обезвоживался в этиловом спирте и заключался в смесь смол эпон–аралдит. Срезы изготавливались на ультратоме «Leica EM UC7» (Австрия). Полутонкие срезы, окрашенные азуром II, изучали в микроскопе «Primo Star» («Carl Zeiss», Германия) с цифровой фотокамерой G-10 («Саnon», Япония) и программным обеспечением «Axio Vision» («Carl Zeiss», Германия). В каждом срезе в 10-ти полях площадью 0,016 мм2 измеряли ядра, ядрышки пинеалоцитов. Ультратонкие срезы, контрастированные уранилацетатом и цитратом свинца, исследовали в трансмиссионном микроскопе «JEM-100 СХ II» (JEOL, Япония). С помощью тест-системы, соответствующей 47 мкм2 среза при увеличении ×25000, в 10-ти случайно отобранных пинеалоцитах измеряли удельный объём органелл. Данные морфометрии обрабатывали с помощью пакета программ «Statistica for Windows», версия 6.1 (StatSoft Inc., США). Для кариометрических показателей вычисляли среднее значение М, стандартное отклонение s, оценивали значимость различий по t-критерию Стьюдента. Для органеллометрических показателей вычисляли медиану Ме, 25%-й и 75%-й квартили. Значимость различий оценивали по U-критерию Манна–Уитни на уровне p<0,05.
Результаты исследования и их обсуждение
У лесной полёвки шишковидная железа имеет целлюлярный тип строения. Пинеалоциты условно подразделяются на светлые (тип I) и тёмные (тип II). Булавовидные отростки пинеалоцитов, транспортирующие секреторные везикулы, обнаруживаются в перикапиллярном пространстве. Капилляры с фенестрированным эндотелием. Вокруг капилляров и между клетками паренхимы выявляются «вакуоли» (рис. 1а).
Среди светлых пинеалоцитов выделяются клетки с комплексами гранулярной эндоплазматической сети (тип Iа) и развитым комплексом Гольджи (тип Ib). Для Ib-клеток характерна нейрокринная секреция. В пинеалоцитах типа Iа находит место нейрокринная и эпендимоподобная секреция.
У совместно содержавшихся сеголеток красной полёвки среди Ib-пинеалоцитов обнаруживаются гипертрофированные клетки. Гипертрофию и гиперплазию претерпевают митохондрии и комплекс Гольджи в их цитоплазме. С активизацией нейросекреции пинеальных гормонов, возможно, антигонадотропного пептида и мелатонина, по-видимому, связано сдерживание полового созревания. Образуются поля синаптических сфер, что также свидетельствует об активизации деятельности железы. В Iа-клетках вокруг крупных комплексов цистерн гранулярной эндоплазматической сети выявляются секреторные везикулы и многочисленные митохондрии. Митохондриальные кристы широкие, матрикс умеренно электронно-плотный (рис. 1б).
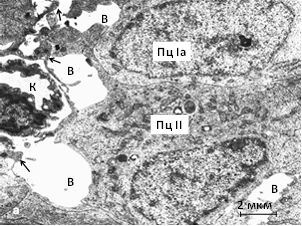
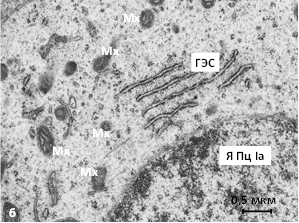
Рис. 1. Ультраструктура шишковидной железы красной полёвки: а – позднелетняя генерация, индивидуальное содержание, б – весенне-летняя генерация, совместное содержание. В – «вакуоли», К – капилляр, Пц – пинеалоциты типа Iа и II, стрелки – булавовидные отростки, ГЭС – гранулярная эндоплазматическая сеть, Мх – митохондрии, Я – ядро. Электронограммы
При совместном содержании ядра Ib-пинеалоцитов у красной полёвки увеличивается на 7,5%, на 27% – удельный объём гранулярной эндоплазматической сети, на 41% – комплекса Гольджи, на 49% – митохондрий. Диаметр ядрышек уменьшается на 5,2%. У красно-серой полёвки сдвиги показателей менее выражены: удельный объём гранулярной эндоплазматической сети уменьшается на 19%, митохондрий – увеличивается на 21% (табл. 1).
Таблица 1
Различия при перенаселении кариометрических показателей (M±s) и органеллометрических (Ме (Q1-Q3), р<0,05) пинеалоцитов типа Ib у лесной полёвки
|
Вид |
красная полёвка |
красно-серая полёвка |
||
|
Содержание |
совместное (n=6) |
индивидуальное (n=4) |
совместное (n=5) |
индивидуальное (n=3) |
|
SЯ, мкм2 |
57,4±11,1* p<0,001 |
53,4±5,4
|
53,4±11,6 |
54,4±12,3 |
|
dЯк, мкм |
1,46±0,23* p<0,001 |
1,54±0,15
|
1,51±0,20
|
1,56±0,15
|
|
VГЭС, % |
3,39* (3,14-3,81) |
2,67 (2,08-3,56) |
1,57* (1,23-2,10) |
1,94 (1,36-2,84) |
|
VМх, % |
8,23* (7,48-8,62) |
5,52 (5,21-6,56) |
7,66* (7,31-7,90) |
6,33 (6,02-7,23) |
|
VКГ, % |
4,44* (4,17-4,72) |
3,14 (2,94-3,59) |
3,08 (2,31-3,88) |
3,47 (3,32-3,66) |
Примечание: * – значимые различия в пределах вида, SЯ – площадь ядра, dЯк – диаметр ядрышка, VГЭС – удельный объём гранулярной эндоплазматической сети в цитоплазме, VМх – митохондрий, VКГ – комплекса Гольджи.
Не исключено, что склонные к эпендимоподобной секреции пинеалоциты с комплексами параллельных цистерн гранулярной эндоплазматической сети являются «добавочными», включаются в гормонпродуцирующую деятельность при стрессе, усиливая нейрокринный эффект «главных» светлых пинеалоцитов со стопками Гольджи.
При совместном содержании у красной полёвки разрастается астроглия, уменьшается число пинеалоцитов, чаще обнаруживаются тёмные, дегенерирующие и гипертрофированные светлые пинеалоциты, а также гигантские межклеточные «вакуоли» с конкрециями. У красно-серой полёвки клеточная гипертрофия не выражена. «Вакуоли» мелкие. Тёмных пинеалоцитов больше в поверхностной зоне органа, чем в глубокой, где выявляются очаги атрофии паренхимы (рис. 2).
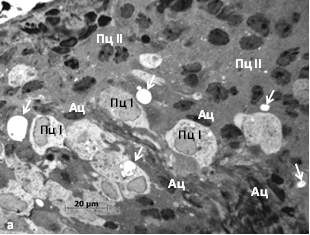
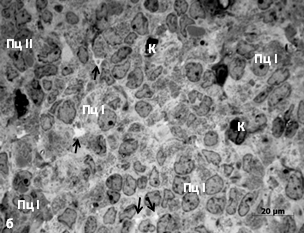
Рис. 2. Шишковидная железа сеголеток позднелетней генерации совместного содержания: а – красная полёвка, поверхностная зона, б – красно-серая полёвка, глубокая зона. Ац – астроциты, К – капилляр, Пц – пинеалоциты типа I и II. Стрелки – межклеточные «вакуоли» с песчинками. Окраска азуром II
Увеличение у красной полёвки числа несекретирующих тёмных пинеалоцитов, гипертрофия светлых свидетельствуют о том, что, в железе, наряду с «подготовкой» к переходу в состояние зимнего «покоя», проявляется активная гормонпродуцирующая деятельность. Утилизация и замещение астроцитами апоптотически гибнущих дегенерирующих пинеалоцитов, возможно, являются отражением подавления другой онтогенетической программы, связанной с быстрым ростом и половым созреванием [9]. Клеточная гипертрофия, формирование гигантских «вакуолей» в местах гибели пинеалоцитов и массового залегания тёмных клеток с небольшими зазубренными ядрами может рассматриваться как проявление приспособительной реакции на стресс. Красную полёвку можно охарактеризовать как вид со стрессорным типом функциональной организации шишковидной железы.
Единичные межклеточные «вакуоли» с конкрециями, наличие пинеалоцитов с узкими перикарионами, бедными органеллами, и небольшими причудливой формы ядрами, очагов атрофии у красно-серой полёвки свидетельствуют о слабой вовлечённости шишковидной железы данного вида в реакцию на стресс. Очевидно, эффект влияния света на циркадианные пейсмекеры у красно-серой полёвки превалирует над экстремальным воздействием высокой плотности популяции. Гипоталамо-гипофизарная, симпатико-адреналовая и половая системы не проявляют такой активности как у красной полёвки. Участие в размножении сеголеток позднелетней генерации у красно-серой полёвки определяется не столько плотностью популяции, как укорочением суточной фотофазы.
У сеголеток красно-серой полёвки позднелетней генерации индивидуального содержания по сравнению с весенне-летней обнаруживается большее число «вакуолей» (рис. 3).
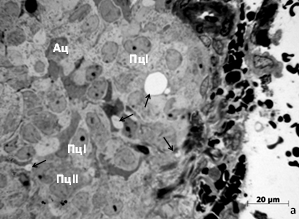
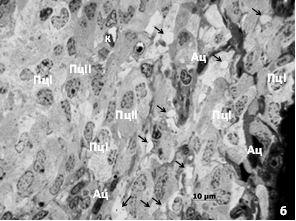
Рис. 3. Участок поверхностной зоны шишковидной железы сеголеток красно-серой полёвки, содержавшихся индивидуально: а – весенне-летняя генерация, б – позднелетняя генерация. Ац – астроциты, стрелки – межклеточные «вакуоли», К – капилляр, Пц – пинеалоциты типа I и II. Окраска азуром II
Карио- и органеллометрические показатели функциональной активности пинеалоцитов у сеголеток лесной полёвки проявляют межвидовой сдвиг с укорочением светлого времени суток. Так, ядра светлых клеток типа Ib красно-серой полёвки весенне-летней генерации на 18%, диаметр ядрышек на 7% больше красной. Удельный объём гранулярной эндоплазматической сети в цитоплазме в 2,4 раза уступает красной. При более короткой суточной фотофазе ядра светлых пинеалоцитов красно-серой полёвки на 28% превосходят площадь их среза у красной полёвки, диаметр ядрышек – на 12%, удельный объём комплекса Гольджи – на 23% больше, гранулярной эндоплазматической сети – в 1,9 раза меньше красной полёвки. Более выраженный сдвиг показателей, отражающих функциональную активность шишковидной железы красно-серой полёвки, возможно, предопределяет задержку их полового созревания, что не всегда проявляется у красной полёвки (табл. 2).
Таблица 2
Кариометрические (M±s) и органеллометрические (Ме (Q1-Q3), р<0,05) показатели
пинеалоцитов типа Ib у лесной полёвки
|
Генерация |
весенне-летняя |
позднелетняя |
||
|
Вид |
красная полёвка (n=6) |
красно-серая полёвка (n=4) |
красная полёвка (n=4) |
красно-серая полёвка (n=4) |
|
SЯ, мкм2 |
54,9±8,3
|
64,8±7,0* p<0,001 |
41,9±10,0 |
53,7±11,8* p<0,001 |
|
dЯк, мкм |
1,51±0,19
|
1,61±0,17* p<0,001 |
1,36±0,21
|
1,52±0,19* p<0,001 |
|
VГЭС, % |
3,32 (2,89-3,66) |
1,36* (0,96-2,40) |
3,12 (2,70-3,26) |
1,63* (1,32-2,40) |
|
VМх, % |
7,32 (6,42-8,39) |
7,46 (6,92-7,84) |
7,18 (6,63-7,59) |
7,26 (6,33-7,66) |
|
VКГ, % |
4,05 (3,41-4,56) |
3,93 (3,12-4,72) |
2,82 (2,08-3,94) |
3,47* (2,80-3,72) |
Примечание: * – значимые различия с красной полёвкой в пределах генерации.
Заключение
Таким образом, на основании проведённого анализа изменений морфофункционального состояния шишковидной железы у двухмесячных самцов-сеголеток двух видов лесной полёвки весенне-летней и позднелетней генераций индивидуального и совместного содержания можно объяснить видовые особенности реализации программы онтогенеза. Программа сдерживания полового созревания у красно-серой полёвки реализуется, благодаря фотозависимому характеру морфофункциональной организации шишковидной железы, на секреторную активность которой выраженное воздействие оказывает продолжительность светлого времени суток. Красная полёвка имеет стрессорный тип функциональной организации шишковидной железы, в большей мере реагирующий не на изменения продолжительности светлого времени суток, а на высокую плотность популяции.
Рецензенты:
Пурлик И.Л., д.м.н., профессор кафедры патологической анатомии ГБОУ ВПО «Сибирский государственный медицинский университет» Министерства здравоохранения РФ, г. Томск;
Мустафина Л.Р., д.м.н., профессор кафедры гистологии, эмбриологии и цитологии ГБОУВПО «Сибирский государственный медицинский университет» Министерства здравоохранения РФ, г. Томск.
Библиографическая ссылка
Герасимов А.В., Костюченко В.П., Денисов А.А. МОРФОФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ ШИШКОВИДНОЙ ЖЕЛЕЗЫ РАЗЛИЧНЫХ ВИДОВ ЛЕСНОЙ ПОЛЁВКИ ПРИ УКОРОЧЕНИИ СВЕТЛОГО ВРЕМЕНИ СУТОК И ВЫСОКОЙ ПЛОТНОСТИ ПОПУЛЯЦИИ // Современные проблемы науки и образования. 2015. № 3. ;URL: https://science-education.ru/en/article/view?id=19950 (дата обращения: 01.07.2026).