


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
IMMUNOHISTOCHEMICAL RESEARCHES OF THYMIC CELLULAR COMPOSITION AT CARCINOGENESIS IN CONDITION OF CONGENITAL IMMUNODEFICIENCY

С точки зрения иммунологии беременность является уникальным состоянием, при котором в организме матери развивается наполовину чужеродный организм. Иммунная система матери проявляет толерантность к антигенным структурам плода, благодаря чему не происходит его отторжения [10]. Механизм этого процесса до сих пор остается неясным, однако продолжают появляться гипотезы, объясняющие этот феномен.
Онтогенез иммунной системы начинается в раннем эмбриональном периоде, продолжается в течение всей внутриутробной жизни и полностью завершается лишь через несколько лет после рождения. Известно, что во внутриутробном и неонатальном периодах плод, а впоследствии – новорожденный приобретает пассивный иммунитет от матери. Через плаценту к плоду поступают антитела, продуцируемые В-лимфоцитами матери. При нормально функционирующей плаценте в крови плода обнаруживаются только материнские иммуноглобулины G, однако при патологиях плаценты возможно попадание в кровь плода и других групп иммуноглобулинов. После рождения факторы иммунитета продолжают поступать в организм новорожденного вместе с материнским молоком. Известно, что в состав грудного молока входят иммуноглобулины, множество цито- и хемокинов, факторы врожденного иммунитета (комплемент, лизоцим, фактор хемотаксиса и др.), пребиотики, гормоны и даже некоторые виды клеток (макрофаги, нейтрофилы, лимфоциты) [9].
Состояние здоровья матери во время беременности также влияет на развитие иммунной системы плода. В современной литературе имеются данные о том, что поражение гепатобилиарной системы матери различной этиологии сопровождается угнетением Т-лимфоцитопоэза у потомства и приводит к нарушению клеточного иммунного ответа в раннем постнатальном периоде. Патология печени матери также ведет к изменению морфофункционального состояния клеток системы мононуклеарных фагоцитов потомства [7]. Длительная прегравидарная алкоголизация матери способствует развитию патоморфологических преобразований паренхимы тимуса новорожденных крыс [5]. Осложнения беременности (острый гестоз, угроза прерывания) являются факторами риска развития у детей тимомегалии [6].
Однако в известных нам источниках литературы мы не обнаружили информации о влиянии иммунодефицитных состояний во время беременности на формирование иммунной системы потомства. Широкая распространенность патологий иммунной системы в последние годы делает эту тему чрезвычайно актуальной. Известно, что у лиц с различными формами иммунодефицитов отмечается повышенная заболеваемость злокачественными новообразованиями [8]. Неуклонный рост онкопатологии, в т.ч. у детей, побуждает поднять вопрос о механизмах канцерогенеза на фоне иммунологической недостаточности.
Цель исследования – изучить влияние иммунодефицитного состояния во время беременности на формирование противоопухолевого иммунитета потомства.
Материал и методы исследования
Эксперименты выполнены на 150 белых нелинейных крысах-самцах в возрасте 3 месяцев. Животные были получены посредством спаривания 4-месячных крыс-самок с самцами массой 150-200 г. Спаривание спленэктомированных самок с самцами проводилось спустя месяц после спленэктомии. Кормление, уход и выведение из эксперимента крыс проводились согласно правилам обращения с лабораторными животными. В качестве модели вторичного иммунодефицита выбрана спленэктомия.
Животные были разделены на четыре группы: 1 – потомство здоровых самок – 30 крыс; 2 – потомство здоровых самок, которым через 1 месяц после рождения внутрибрюшинно вводили канцероген (1,2-диметилгидразин) из расчета 20 мг/кг 1 раз в неделю в течение 4 недель (общая доза составила 80 мг/кг) – 40 крыс; 3 – потомство спленэктомированных самок – 40 крыс; 4 – потомство спленэктомированных самок, которым через 1 месяц после рождения вводили канцероген по той же схеме – 40 крыс.
Объектом исследования служил тимус. При патоморфологическом исследовании учитывались частота развития новообразований, их морфологические особенности, локализация.
Для исследования срезов тимуса использовали следующие методы.
1. Окраска гематоксилином и эозином с последующей морфометрией коркового и мозгового вещества долек.
2. Морфометрический метод с использованием программы «Микро-Анализ» для измерения толщины коркового и площади мозгового вещества тимуса, оценки иммуногистохимических реакций.
3. Иммуногистохимические методы с использованием моноклональных (МКАТ) и поликлональных (ПКАТ) антител фирмы Santa Cruze (США) к крысиным антигенам:
а) ПКАТ к CD3 – для идентификации зрелых тимоцитов дольки;
б) МКАТ к CD68 – маркер макрофагов в структурах микроокружения долек тимуса;
в) МКАТ к белку S100 – маркеру антигенпрезентирующих дендритных клеток;
г) ПКАТ к синаптофизину – для идентификации клеток нейроэндокринного происхождения, к которым в тимусе относятся клетки APUD-серии;
д) МКАТ к панцитокератину – для неселективной идентификации эпителиальных клеток дольки.
4. Статистическая обработка полученных цифровых данных проведена с помощью пакета программ Microsoft office (Word и Excel). В работе приводятся следующие показатели: М — средняя арифметическая величина; m — средняя ошибка средней арифметической величины. Статистическую достоверность оценивали по t-критерию Стьюдента (t).
Результаты исследования и обсуждение
Масса тимуса потомства интактных крыс в возрасте 3-х месяцев составляет 304,2±24,0 мг. На окрашенных гематоксилином и эозином срезах хорошо определяются дольки округлой или полигональной формы со светлым мозговым и темным корковым веществом. Толщина коркового вещества равна 341,2±14,7 мкм, площадь мозгового вещества – 0,43±0,076 мм2.
При проведении иммуногистохимической реакции на CD3 для выявления зрелых Т-клеток обнаружено, что они расположены преимущественно в мозговом веществе долек и занимают 38,3±1,9% площади. В корковом веществе CD3+ клетки также встречаются, но занимают лишь 13,7±0,7% площади (рис. 1).
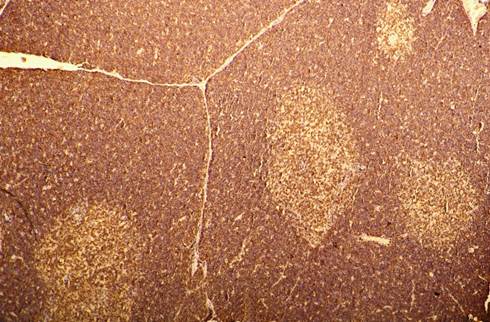
Рис. 1. Тимус потомства интактной крысы. Экспрессия CD3 в тимоцитах дольки. Иммуногистохимическая реакция с CD3, ув. 100x.
Установлено, что CD68+ клетки в тимусе потомства интактных крыс расположены в один ряд вокруг мозгового вещества, их количество составляет 17,5±1,4 клетки в поле зрения. Единичные макрофаги также встречаются и в корковом веществе долек. S100+ (дендритные) клетки главным образом встречаются в области кортико-медуллярной границы одиночно или группами по 2-3 клетки.
Положительная реакция на ПКАТ к синаптофизину обнаруживается у единичных клеток в корковом веществе. Панцитокератин-позитивные клетки (эпителиоциты) расположены равномерно по всей площади дольки и имеют сетевую организацию. В промежутках между эпителиальными клетками находятся тимоциты. Эпителиоциты мозгового вещества имеют более крупные размеры и занимают 31,6±1,1% площади. Эпителиальные клетки коркового вещества меньше мозговых и занимают 28,1±0,9% площади.
Нами выявлено, что беременность на фоне иммунодефицита и последующее введение канцерогена потомству приводит к выраженным морфологическим изменениям в тимусе уже через 30 суток после окончания курса инъекций, т.е. в возрасте 3-х месяцев. Установлено, что масса тимуса уменьшается во всех опытных группах по сравнению с контрольной и в группе потомства спленэктомированных крыс с введением канцерогена составляет 293,0±7,2 мг. Наиболее выраженные изменения массы наблюдаются в тимусе потомства спленэктомированных крыс без постнатального воздействия – показатель равен 236,2±30,9 мг (p<0,001).
Изменения наблюдаются и при морфометрии срезов, окрашенных гематоксилином и эозином. Так, в дольках тимуса потомства спленэктомированных самок площадь мозгового вещества уменьшается в 1,7 раза и равна 0,16±0,027 мм2. Толщина коркового вещества изменяется незначительно и составляет 300,7±14,2 мкм. В группе потомства спленэктомированных самок с введением канцерогена, напротив, большим изменениям подвержена толщина коркового вещества – она уменьшается в 2 раза по сравнению с контрольной группой и составляет 152,7±6,3 мкм. Площадь мозгового вещества уменьшается на 23% и равна 0,21±0,024 мм2 (p<0,01). Дольки деформированы, наблюдается смещение мозгового вещества на периферию, границы между дольками стерты.
Результаты подсчёта иммуногистохимических реакций в тимусе потомства спленэктомированных самок отличаются от аналогичных исследований контрольной группы. Особо значимо увеличивается численность макрофагов (в 1,5 раза) и клеток APUD-серии (более чем в 3 раза). Медуллярные эпителиоциты расходятся к периферии мозгового вещества, при этом в центральной области эпителиальные клетки отсутствуют (рис. 2).
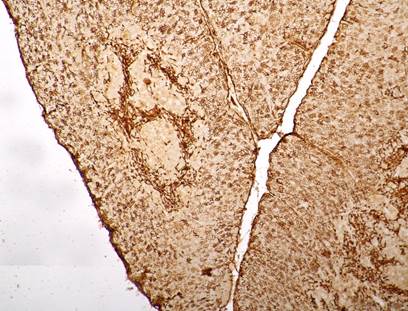
Рис. 2. Тимус потомства спленэктомированных самок. Смещение медуллярных эпителиоцитов к периферии мозгового вещества. Иммуногистохимическая реакция с Pan-cytokeratin, ув. 100х.
В тимусе потомства интактных крыс с последующим введением канцерогена наблюдается уменьшение количества CD3+ клеток как в корковом, так и в мозговом веществе, причем более значительное уменьшение (на 13%) в мозговом веществе (p<0,001). Количество S100+ и синаптофизин+ клеток, напротив, увеличивается. Клетки APUD-серии концентрируются на границе кортико-медуллярной зоны, а также встречаются в мозговом веществе (5-7 в поле зрения). Количество клеток, экспрессирующих CD68, а также корковых эпителиоцитов изменяется недостоверно, в то время как медуллярные эпителиальные клетки смещаются в область кортико-медуллярной границы.
В тимусе потомства спленэктомированных крыс с последующим введением канцерогена наблюдается еще более значительное уменьшение количества CD3+. В мозговом веществе они занимают лишь 20,2±2,1% площади (p<0,001). Количество макрофагов, напротив, увеличивается. Они располагаются в 2-3 ряда в премедуллярной зоне, а также в мозговом веществе долек. Количество синаптофизин+ и S100+ клеток также возрастает. Клетки, экспрессирующие панцитокератин, - эпителиоциты - в мозговом веществе располагаются плотной сетью на границе коркового и мозгового вещества, в центре мозгового вещества они отсутствуют, в то время как корковые эпителиоциты сохраняют сетевую организацию.
Заключение
Таким образом, нами выявлено, что удаление селезенки у самок влияет на морфофункциональное состояние тимуса их потомства. Это выражается в снижении его массы, уменьшении толщины коркового и площади мозгового вещества, увеличении количества макрофагов и клеток APUD-серии, изменении расположения медуллярных эпителиоцитов. Введение канцерогена потомству спленэктомированных самок вызывает более выраженные инволютивные изменения в тимусе по сравнению с введением канцерогена потомству интактных крыс: уменьшается количество CD3+ клеток с одновременным увеличением числа CD68+, S100+ и синаптофизин+ клеток, медуллярные эпителиальные клетки смещаются в премедуллярную зону.
Механизмы развития акцидентальной инволюции тимуса на фоне развития опухоли до сих пор остаются до конца не выясненными. Одни авторы считают пусковым фактором прямую индукцию апоптоза тимоцитов и уменьшение процента Т-клеток в S-стадии клеточного цикла. Другие считают наиболее значимым недостаточное поступление в тимус клеток-предшественников. Причём в костном мозге они сохраняются в достаточном количестве и являются функционально полноценными. Вероятно, это может быть следствием миграции клеток-предшественников из костного мозга в селезенку или опухоль [2]. Существует также мнение, что акцидентальная инволюция тимуса развивается как адаптивная реакция на стресс любой этиологии. Индукторами инволюции могут выступать глюкортикоидные гормоны, а также такие цитокины как TNF-α, IL-1, IL-4, TGF-β, VEGF [4].
Мы считаем, что инволютивные процессы в тимусе развиваются как вследствие токсического влияния самой опухоли [3], так и по причине дисфункции взаимодействия в системе «надпочечники-гипофиз-тимус» [1]. Посредниками взаимодействия эндокринной и иммунной систем в этом случае являются дендритные клетки, способные при их стимуляции секретировать те или иные иммунорегулирующие факторы.
Рецензенты:
Гунин А.Г., д.м.н., профессор, профессор кафедры акушерства и гинекологии ФГБОУ ВПО «Чувашский государственный университет им. И.Н. Ульянова», г. Чебоксары.
Смелов С.В., д.м.н., доцент кафедры нормальной и топографической анатомии с оперативной хирургией ФГБОУ ВПО «Чувашский государственный университет им. И.Н. Ульянова», г. Чебоксары.
Библиографическая ссылка
Драндрова Е.Г., Стручко Г.Ю., Меркулова Л.М., Кострова О.Ю., Москвичев Е.В., Михайлова М.Н., Арлашкина О.М. ИММУНОГИСТОХИМИЧЕСКИЕ ИССЛЕДОВАНИЯ КЛЕТОЧНОГО СОСТАВА ТИМУСА ПРИ КАНЦЕРОГЕНЕЗЕ В УСЛОВИЯХ ВРОЖДЕННОГО ИММУНОДЕФИЦИТА // Современные проблемы науки и образования. 2015. № 3. ;URL: https://science-education.ru/en/article/view?id=19825 (дата обращения: 30.07.2026).