


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE INFLUENCE OF WATER SALINITY ON THE GROWTH AND REPRODUCTION OF WATER HYACINTH AT NUTRIENT LOAD

Водные макрофиты способны интенсивно поглощать и накапливать многие минеральные и органические вещества, входящие в состав промышленных и хозяйственно-бытовых сточных вод. Ассимиляционная способность гидрофитов широко используется для биоремедиации промышленно загрязненных вод [1-3, 6-10, 14].
В последнее время подробно исследованы процессы поглощения, ассимиляции и детоксикации водными макрофитами тяжелых металлов, фенолов, нефтепродуктов, СПАВ и др. [1, 2, 7-10, 12-14]. Однако исследованиям солеустойчивости и деминерализационной способности гидрофитов уделяется мало внимания [4, 5, 15]. Определение верхних порогов и изучение адаптационных механизмов солеустойчивости гидрофитов является важной задачей в связи с оптимизацией подбора видов и конструирования биоплато для фиторемедиации природных и сточных вод в аридных зонах и при комбинированном загрязнении [5, 7].
Наиболее широкое применение в фиторемедиации комплексно загрязненных сточных вод и природных водоемов имеет водный гиацинт (Eichornia crassipes) – представитель семейства понтедериевых – южноамериканский интродуцент. Это многолетнее, плавающее в поверхностном слое воды розеточное растение (плейстонный гидрофит) – доминант небольших водоемов и водотоков тропической и субтропической зон.
Экологические особенности водного гиацинта следующие [3, 15–18]:
- светолюбивый;
-теплолюбивый (рост и размножение: +16-33°С, оптимум: +25-29°С, гибель: +4°С);
- размножение вегетативное с очень высокой скоростью (до 70 раметов в месяц);
- способность расти и размножаться в среде, содержащей высокие концентрации загрязняющих веществ, вызывающие гибель других водных растений;
- высокая скорость поглощения и биоаккумуляции токсикантов.
Установлен выраженный аллелопатический эффект сырого экстракта эйхорнии по отношению к зеленым водорослям Chlorella vulgaris и Dictyochloropsis splendida и к цианобактериям Spirulina platensis, Nostoc piscinale. Наибольшее подавление роста и размножения наблюдается у Chlorella vulgaris [17].
Имеются фрагментарные сведения о влиянии солёности воды на рост и размножение водного гиацинта [15, 16, 18]. Холм и др. отмечали, что водной гиацинт не выносит солоноватой воды [16]. Экспериментально определена обратная зависимость между продуктивностью E. crassipes и минерализацией воды [18]. При солености выше 6 ‰ прекращается рост, образуются некротические пятна на листьях, а выше 8 ‰ – необратимые физиологические повреждения [15, 18].
Целью работы является изучение повышенной солености воды (2 ‰, 4 ‰ и 6 ‰) в условиях стартовой азотной и фосфорной нагрузки на рост, развитие и размножение лабораторной культуры водного гиацинта.
Материал и методы исследования
Исследования накопительной культуры водного гиацинта проводили на базе отдела биотехнологий и фиторесурсов Института биологии южных морей (г. Севастополь) в условиях лабораторного эксперимента при круглосуточном освещении лампами DRLS в 4-х бассейнах объемом 100 л. В качестве питательной среды использовали водные вытяжки низинного торфа (10 кг торфа / 100 л воды). После загрузки торфа и заполнения бассейнов пресной водой провели отстаивание в течение 3 суток. Объем водной среды составлял 50,0 л. На 3-и сутки к объему среды добавили навески морской соли для получения растворов 2 ‰, 4 ‰ и 6 ‰. Для контроля использовали пресную воду с минерализацией 0,5 ‰. Уровень воды в бассейне поддерживали, компенсируя испарение воды добавлением профильтрованной водопроводной воды.
В качестве имитации залповой эвтрофикации водоема произведено однократное внесение в бассейны по 50 г карбамида и 20 г К2НРО4. В результате превышение ПДК по нормативам для рыбохозяйственных водоемов эвтрофного типа составило для мочевины – 12,5 раз, фосфатов – 2 раза [11]. Активная реакция среды – 9,0. Среду нейтрализовали добавлением 0,1 % раствора соляной кислоты под контролем буферного раствора соды.
Таблица 1
Условия проведения эксперимента*
|
№ эксперимент группы |
1 |
2 |
3 |
4 |
|
Серия эксперимента |
контроль |
2 ‰ |
4 ‰ |
6 ‰ |
|
Температура воды, °С |
26,0-31,0 |
26,0-30,0 |
26,0-29,0 |
26,0-31,0 |
|
29,1±1,1 |
28,6±0,7 |
28,2±0,9 |
29,3±1,2 |
|
|
Активная реакция, лог.ед. |
7,31-8,53 |
6,50-7,67 |
6,30-8,00 |
6,50-8,20 |
|
7,98±0,35 |
7,00±0,30 |
7,00±0,42 |
7,52±0,37 |
* над чертой – пределы колебаний, под чертой – среднее и среднеквадратичное отклонение.
Продолжительность эксперимента – 55 суток (17.02-14.04.2014). Интенсивность освещения у листовой поверхности (3000 лк) контролировали с помощью люксметра Ю-116. Активную реакцию среды измеряли с помощью иономера И-160М.
Ежедневно в 11 ч. в бассейнах измеряли температур и рН воды. Температуру воды регулировали с помощью аквариумных обогревателей AQ-200W. Температуру воздуха поддерживали в пределах: 19–25 °С.
Биомассу определяли в начале эксперимента и, начиная с 22 суток, через каждые 3 дня. В течение первых 22 суток стресса растения не извлекали. Для взвешивания растения кратковременно подсушивали между двумя слоями фильтровальной бумаги. Измеряли морфометрические параметры (длину черешка и корней, длину и ширину листовой пластинки), количество раметов, проводили взвешивание каждого растения с помощью аналитических весов ВЛР-200 с пределами измерений 1 мг – 200 г по ГОСТ 24104-80. Затем растения возвращали в культивационные бассейны. Сырую биомассу пересчитывали на абсолютно сухое вещество (АСВ) по коэффициенту 0,088.
Результаты исследования и их обсуждение
В период стресса, вызванного избыточной биогенной нагрузкой, отмечалось развитие хлороза, некротических пятен и краевого усыхания листьев, а также отмирание части корней и их замещение новыми. Динамика биомассы имеет фазовый характер (рис. 1).
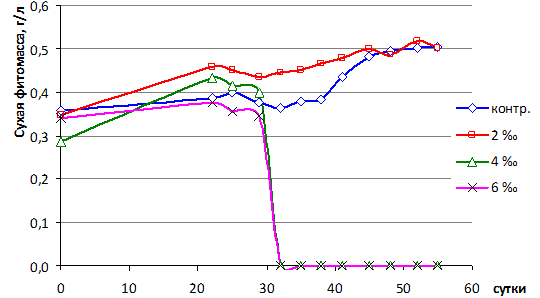
Рис. 1. Динамика биомассы (г/л сухого вещества) водного гиацинта в бассейнах с разной соленостью среды
В период первых 22 суток прирост биомассы отмечался при солености 2 ‰ и 4 ‰, продуктивность была сравнительно высокой за счет активного роста корней. В контроле и при солености 6 ‰ биомасса значимо не изменялась (табл. 2).
Таблица 2
Динамика биомассы (B, мг/л АСВ) и суточной продукции (Pn, мг/л сут. АСВ) при разной солености среды
|
Количество суток |
контроль |
2,0 ‰ |
4,0 ‰ |
6,0 ‰ |
||||
|
B |
Pn |
B |
Pn |
B |
Pn |
B |
Pn |
|
|
0 |
357,3 |
1,4 |
348,5 |
5,0 |
286,9 |
6,7 |
339,7 |
1,6 |
|
22 |
387,2 |
458,5 |
433,8 |
375,8 |
||||
|
25 |
400,4 |
4,4 |
450,0 |
-2,8 |
413,6 |
-6,7 |
355,5 |
-6,7 |
|
29 |
374,9 |
-6,4 |
434,7 |
-3,8 |
398,6 |
-3,7 |
345,0 |
-2,6 |
|
32 |
364,1 |
-3,6 |
446,7 |
4,0 |
0,0 |
0 |
0,0 |
0 |
|
35 |
378,9 |
4,9 |
449,9 |
1,1 |
0,0 |
0 |
0,0 |
0 |
|
38 |
383,9 |
1,6 |
465,2 |
5,1 |
0,0 |
0 |
0,0 |
0 |
|
41 |
434,9 |
17,0 |
480,0 |
4,9 |
0,0 |
0 |
0,0 |
0 |
|
45 |
482,2 |
11,8 |
499,8 |
5,0 |
0,0 |
0 |
0,0 |
0 |
|
48 |
493,7 |
3,8 |
485,8 |
-4,7 |
0,0 |
0 |
0,0 |
0 |
|
52 |
503,0 |
2,3 |
518,7 |
8,2 |
0,0 |
0 |
0,0 |
0 |
|
55 |
505,1 |
0,7 |
501,8 |
-5,6 |
0,0 |
0 |
0,0 |
0 |
Период – 22–32 сутки был критическим, происходило отмирание поврежденных и медленный рост молодых листьев. В контроле и в среде 2 ‰ образовались 1 и 2 рамета (рис. 2). В опытах с 4 и 6 ‰ развития молодых листьев и размножения не произошло, растения погибли. Прирост биомассы не компенсировал потери некромассы, поэтому в целом баланс был отрицательным и лишь в контроле – положительным (табл. 2).
Период прироста биомассы в результате формирования и развития молодых раметов начался с 33-х суток в контроле и с 30-х сут. в 2 ‰ среде и продолжался до 45 суток. Наибольшая удельная суточная продуктивность (P/B) – 4,4 % сут-1 отмечалась в контроле в 38–41 сутки. При солености 2 ‰ наблюдался медленный рост. Высокая скорость размножения отмечалась в период 41–52 сутки в контроле (максимум: 48–52 сутки – 1,3 рамета/сут.), а в 2 ‰ среде она была значительно ниже (максимум: 45–48 сутки – 0,33 рамета/сут.). Различия по критерию Крускала-Уоллеса значимые (H=4,05, p=0,044).
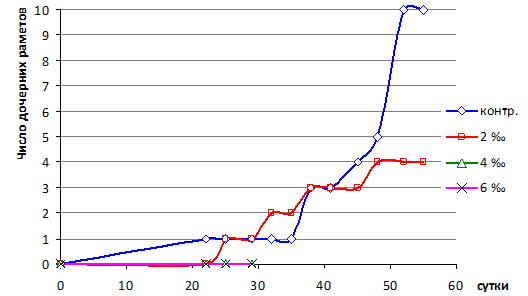
Рис. 2. Динамика образования раметов водного гиацинта в бассейнах с разной соленостью среды
В период – 45–55 сутки наряду с ростом и размножением началось усыхание поврежденных листьев, наиболее выраженное при 2 ‰ солености. В период – 52–55 сутки рост и размножение прекратились (рис. 2), что вызвано исчерпанием биогенных элементов в воде.
Множественное последовательное сравнение биомасс всех серий опыта с контрольной по критерию Крускала-Уоллеса показало статистически значимые различия по биомассе (K=31, p=0,0001) и скорости размножения (K=34, p=0,0001). Парное сравнение критерием Дана (p<0,05) выявило значимые различия по этим показателям между контролем и группами 4 ‰, 6 ‰, а также между группами 2 ‰ и 4 ‰; 2 ‰ и 6 ‰.
Корреляционный анализ динамики биомассы и суточной продукции критерием ранговой корреляции Спирмена показал следующее (табл. 3).
Таблица 3
Ранговая корреляция биомассы и чистой продукции при разной солености
|
Корреляция Спирмена |
Биомасса |
Суточная продукция |
||||
|
2,0 ‰ |
4,0 ‰ |
6,0 ‰ |
2,0 ‰ |
4,0 ‰ |
6,0 ‰ |
|
|
контроль |
0,951 |
-0,164 |
-0,164 |
0,230 |
0,214 |
0,214 |
|
2,0 ‰ |
|
-0,294 |
-0,294 |
|
0,518 |
0,518 |
|
4,0 ‰ |
|
|
1,000 |
|
|
1,000 |
Жирным шрифтом выделены значимые корреляции показателей (P<0,05).
Наибольшую согласованность (r=1,00; p=0,0001) имеет динамика биомассы и суточной продукции при солености 4 и 6 ‰. Высокая корреляция (r=0,95 p=0,0001) отмечена по биомассе в контроле и в среде 2 ‰. Однако в этой группе сравнения суточная продукция значимо не коррелированна (r=0,23 p=0,485), что объясняется различием в периодах размножения и отмирания старых листьев.
Динамика биомассы коррелирует с изменением pH среды: r=0,81, p=0,002 (контроль) и r=0,97, p=0,0001 (2 ‰ раствор). Активный рост и увеличение биомассы гиацинта вызывают смещение pH среды в щелочную сторону, что мы наблюдаем в ходе эксперимента (рис. 3).
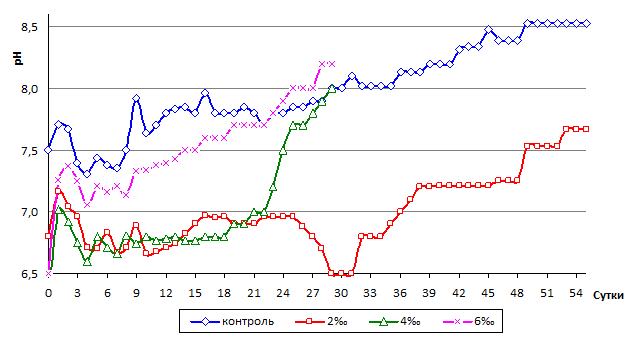
Рис. 3. Динамика pH в культивационных бассейнах с разной соленостью среды
Выводы
1. Изменения биомассы в контроле и опытной серии солености среды имеют фазовый характер. В начальный период отмечается прирост биомассы, наиболее выраженный при солености 2 ‰ и 4 ‰, с последующим угнетением и гибелью на 32-е сутки растений в среде 4 ‰, 6 ‰ и восстановлением культуры в 2 ‰ среде и пресной воде (контроль).
2. Активное вегетативное размножение отмечается в контроле в период 41–52 сутки (максимум: 1,3 рамет/сут.). При солености 0,2 % скорость размножения значительно меньше (максимум: 0,33 рамет/сут.).
3. Динамика биомассы коррелирует с изменением активной реакции среды, отражающей увеличение скорость фотосинтеза.
4. Критической соленостью для водного гиацинта в условиях высокой стартовой биогенной нагрузки является 2 ‰.
Рецензенты:
Дмитриев А. И., д.б.н., профессор, заведующий кафедрой биологии, химии и биолого-химического образования ФГБОУ ВО «Нижегородский государственный педагогический университет им. К. Минина», г. Нижний Новгород;
Воскресенская О. Л., д.б.н., профессор, директор института биологии и естественных наук и заведующая кафедрой экологии ФГБОУ ВО «Марийский государственный университет», г. Йошкар-Ола.
Библиографическая ссылка
Макеев И.С., Горбунова С.Ю., Коротаева М.И. ВЛИЯНИЕ СОЛЁНОСТИ ВОДЫ НА РОСТ И РАЗМНОЖЕНИЕ ВОДНОГО ГИАЦИНТА В УСЛОВИЯХ БИОГЕННОЙ НАГРУЗКИ // Современные проблемы науки и образования. 2014. № 6. ;URL: https://science-education.ru/en/article/view?id=17139 (дата обращения: 29.05.2026).