


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
ESTIMATION THE HERITABILITY OF SELECTED BREEDING MATERIAL BY SPEED OF GROWTH OF THE SEED PROGENY IN THE CONDITIONS DRY STEPPE LOWER VOLGA REGION

Изучение влияния генетических факторов и факторов среды на скорость роста основных лесообразователей является одной из актуальнейших проблем лесовосстановления. От того, как соотносятся наследственные и экологические факторы, влияющие на рост дерева, в конечном итоге зависит стратегия лесовосстановления. В частности, от того, насколько сильным является влияние генотипа на рост дерева, зависит целесообразность применения методов селекции для повышения продуктивности древостоев, а также возможность разработки использования методов ранней диагностики быстрорастущих деревьев при создании лесных культур.
В настоящее время среди исследователей нет однозначного мнения относительно степени влияния генотипов на рост и устойчивость деревьев основных лесообразующих пород в защитных насаждениях. Значительная часть исследователей считает, что генотип - решающий фактор роста и устойчивости дерева [1; 2]. Другие исследователи считают влияние генотипа на рост недостоверным [5].
Степень генетической обусловленности роста определяет также возможность разработки и использования методов ранней диагностики деревьев, отличающихся высокой скоростью роста, что является одним из приоритетных направлений.
Влияние генетических факторов на рост и устойчивость можно оценить на основе изучения интенсивности дифференциации деревьев по скорости роста и устойчивости. В частности, пристальное внимание исследователей привлекает возраст стабилизации рангового положения деревьев в насаждении, дающий ориентировочную оценку сроков испытания ценных генотипов. Данные исследователей расходятся в отношении оценки возраста, при котором происходит стабилизация рангов высот саженцев, позволяющая производить сравнительную оценку их роста и, следовательно, предсказывать преобладание в будущем по высоте тех или иных деревьев. Многие исследователи указывают возраст 5-7 лет. По другим данным, ранговое положение отдельных особей стабилизируется и определяется в составе древостоя с 10-15 лет. Есть также данные, что указанная стабилизация роста дерева происходит в 20-30 лет. Кроме того, существует мнение, что полная оценка всех признаков и свойств деревьев с целью дальнейшего размножения лучших из них возможна лишь в 60-100 лет. Видимо, разница в оценке минимального возраста, при котором возможно сравнение роста деревьев, во многом обусловлена различным подходом исследователей к допустимой точности результатов работы [1; 2].
Одним из спорных вопросов защитного лесоразведения является степень снижения генетического разнообразия насаждений при проведении селекционных мероприятий. При использовании улучшенного семенного материала с объектов постоянной лесосеменной базы (прежде всего, с лесосеменных плантаций) проблема сохранения генетического полиморфизма встает особенно остро, поскольку популяции, служащие источником таких семян, характеризуются ограниченных числом генотипов. В связи с этим одним из важнейших условий дальнейшего совершенствования селекционных мероприятий является оценка уровня генетического разнообразия семенного материала объектов постоянной лесосеменной базы [2; 5].
Определение степени генетической обусловленности скорости роста деревьев сосны проводилось на основе оценки различий между семьями с использованием дисперсионного анализа. При этом оценивалась достоверность различий, а также доля аддитивной генетической составляющей в фенотипической изменчивости признака, рассчитываемая как коэффициент наследуемости в узком смысле (h2). При оценке степени генетической обусловленности скорости роста деревьев использовался также метамерно-генетический анализ, позволяющий без смены поколений определить долю генетипической изменчивости в общей фенотипической изменчивости при росте дерева по высоте. При этом определялись генетическая, экологическая (паратипическая) и общая фенотипическая дисперсия приростов, а также отношение генотипической дисперсии к фенотипической, т.е. коэффициент наследуемости в широком смысле (Н2).
Для оценки эффективности фенотипического отбора быстрорастущих деревьев сосны производилось сравнение роста культур, созданных с использованием улучшенных семян (полусибсовое потомство плюсовых деревьев), с ростом культур из семян нормальной селекционной категории (контроль).
Влияние отбора плюсовых деревьев на генетическое разнообразие их потомства определили на основе сравнения генотипического варьирования семенного потомства плюсовых деревьев с соответствующими показателями смеси семян нормальной селекционной категории. При этом рассчитывались генетическая и паратипическая составляющие годичного прироста на основе метамерно-генетического анализа. Рассчитывался также коэффициент наследуемости в широком смысле (Н2), который представляет собой соотношение этих показателей и отражает уровень генетического разнообразия признака.
Проведенные исследования показали, что деревья сосны существенно различаются по скорости роста в высоту. В частности, деление совокупности деревьев на 5 групп роста на основе кластерного анализа демонстрирует дифференциацию деревьев в культурах по скорости роста в высоту. Полученный при помощи кластерного анализа ход роста деревьев разных групп роста свидетельствует о наличии четких различий в средних высотах растений между указанными группами в течение всего периода наблюдений (рис. 1).
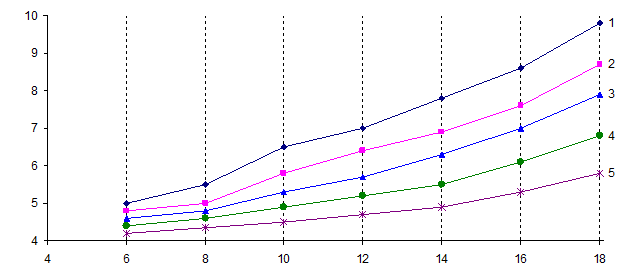
Рис. 1. Изменение высоты сосны разных групп роста в селекционных лесосеменных плантациях Новоаннинского лесхоза: 1 - наиболее быстрорастущие деревья (1-я группа); 2 - деревья ускоренного роста (2-я группа); 3 - деревья со средней скоростью роста (3-я группа); 4 - деревья замедленного роста (4-я группа); 5 - медленно растущие особи (5-я группа).
По данным дисперсионного анализа, достоверные на 5% -ном уровне значимости различия по высоте между деревьями разных групп роста проявляются начиная с 4-летнего возраста. При этом в процессе онтогенеза кривые хода роста расходятся, т.е. дивергенция групп со временем увеличивается (межгрупповое варьирование по высоте возрастает быстрее, чем внутригрупповое), что говорит о возрастании различий по высоте между быстро и медленно растущими деревьями.
Быстрорастущие клоны и семьи, отобранные в 4-летнем возрасте при посадке на лесокультурную площадь, сохраняют преимущества в росте. Это, во-первых, подтверждает возможность ранней диагностики деревьев-лидеров, а во-вторых, свидетельствует, что дифференциация деревьев сосны по скорости роста определяется не условиями микросреды, а индивидуальными генетическими свойствами деревьев.
Для того чтобы установить, в какой мере указанные различия обусловлены генотипом дерева, необходимо провести анализ, позволяющий выявить вклад генетической составляющей в общую дисперсию признака. Принадлежность деревьев на клоновых и семейственных плантациях к той или иной семье (полусибсовому потомству одного дерева), а также относительно выровненные лесорастительные условия участков дают возможность провести такой анализ [2].
Как показали результаты дисперсионного анализа, между семьями существуют достоверные различия (уровень значимости р < 0,01) по основным биометрическим показателям, что говорит о значительном влиянии генетических факторов на их проявление. Расчет значений коэффициентов наследуемости, оценивающих долю общей (коэффициент наследуемости в широком смысле Н2) и аддитивной (коэффициент наследуемости в узком смысле h2) генетических вариаций в фенотипической дисперсии признака для лесосеменных плантаций сосны 20-летнего возраста дал следующие результаты: для высоты растений коэффициент наследуемости в широком смысле, рассчитанный методом метамерно-генетического анализа, составляет в среднем 0,44 (с колебаниями от 0,26 до 0,73). Коэффициент наследуемости в узком смысле высоты растений, рассчитанный на основе дисперсионного анализа, составил 0,14-0,15. Коэффициент наследуемости в узком смысле диаметра ствола составил 0,03-0,10.
Таким образом, исследования показали, что в отличие от ранней диагностики отдельных деревьев сосны, дающей в условиях сухой степи неплохие результаты, отбор быстрорастущих семей в раннем возрасте нецелесообразен потому, что в состав каждой семьи входят деревья, имеющие разные генотипы и различающиеся по скорости роста. Отдельная семья является достаточно гетерогенной смесью, и ранговое положение семьи по среднему значению биометрического показателя в ряду других семей в раннем возрасте достаточно неустойчиво.
Рецензенты:
Васильев Ю.И., д.с.-х.н., профессор, главный научный сотрудник Всероссийского НИИ агролесомелиорации Российской академии наук, г. Волгоград.
Рулев А.С., д.с.-х.н., заместитель директора по науке Всероссийского НИИ агролесомелиорации Российской академии наук, г. Волгоград.
Библиографическая ссылка
Макаров В.М., Иозус А.П., Морозова Е.В. ОЦЕНКА НАСЛЕДУЕМОСТИ ОТОБРАННОГО СЕЛЕКЦИОННОГО МАТЕРИАЛА ПО СКОРОСТИ РОСТА СЕМЕННОГО ПОТОМСТВА В УСЛОВИЯХ СУХОЙ СТЕПИ НИЖНЕГО ПОВОЛЖЬЯ // Современные проблемы науки и образования. 2014. № 4. ;URL: https://science-education.ru/en/article/view?id=14317 (дата обращения: 28.07.2026).