


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
CHANGES ON THE CHLOROPLAST AND MITOCHONDRIAL GENETIC SYSTEMS OF STRAWBERRY BECAUSE OF DROUGHT STRESS

Национальное богатство любой страны, наравне с учитываемыми всеми материальными ценностями, включает землю, недра и зеленый покров. Генетический фонд полезных растений является самым драгоценным богатством страны и народа, к которому должно быть всегда внимательное, заботливое отношение. Это предполагает сбор, накопление растительных ресурсов и его рациональное использование.
Непрерывный рост сельскохозяйственного производства требует внедрение сортов, характеризующихся высокой продуктивностью и адаптационной способностью к действию экстремальных факторов среды. При этом следует отметить что, абсолютная величина устойчивости одного и того же сорта существенно меняется под влиянием разнообразных условий внешней среды, при которых развиваются растения. Поэтому для сопоставления устойчивости разных видов или сортов растений следует ориентироваться не на абсолютную (изменяющуюся в разных циклах оценки), а на их относительную устойчивость (различия уровня устойчивости относительно друг друга), принадлежность их к определенным группам устойчивости [3].
Проблема изучения генетических основ засухоустойчивости у культурных растений, в частности, у ягодных культур, является одной из важнейших задач. Известно что, некоторая часть клеточной ДНК локализована в структурах цитоплазмы - митохондриях и хлоропластах. Здесь протекают процессы репликации, транскрипции и трансляции, обеспечивающие формирование и функционирование этих органоидов.
С историей открытия цитоплазматических и ядерных генетических систем растений тесно связан вопрос об эволюционном происхождении эукариотизма и цитоплазматических генов. Этот вопрос широко дискутировался на протяжении многих лет и обсуждается в настоящее время, что вызвано необходимостью установления общности и специфичности ядерной, хлоропластной и митохондриальной генетических систем высших растений. Особое значение приобретает факт, показывающий, что жизнеспособность организмов, устойчивость к выживанию и многие другие фундаментальные признаки определяются кооперацией различных генетических систем - каждая из них не может отвечать за эти признаки независимо друг от друга. По-видимому, такое взаимодействие вырабатывается в процессе эволюции под влиянием естественного отбора на основе изменчивости генотипа и плазмотипа и обеспечивает координирование всех наследственных систем клетки. Можно предположить, что любая хромосомная мутация способствующая лучшей устойчивости растений к экстремальным факторам только в этом случае будет подхвачена естественным отбором, если индуцируемые ею ферменты благоприятно взаимодействуют с энзимами, кодируемыми митохондриальными и пластидными плазмогенами [1,10]. С другой стороны, митохондриальная мутация, которая обуславливает более быстрое деление клеток, по сравнению с нормальным уровнем, может быть обнаружена, что даёт начало новому сорту только в том случае, если быстрая репродукция клеток будет нормально сочетаться c продуктами хромосомных или пластидных генов. В противоположном случае, такая мутация может разрушить клетку и не проявиться в потомстве. Цитоплазматическая наследственность у растений реализуется через ядерно-цитоплазматические взаимодействия [4,6], т.е. через тесную кооперацию и интеграцию систем генома и плазмона. Следует отметить, что с типом цитоплазмы связаны такие биологические функции растений, как иммунитет и устойчивость к неблагоприятным природным факторам [7,8,9].
В последние годы появились работы, анализирующие молекулярные изменения ДНК и РНК хлоропластов и митохондрий [2]. Эти данные позволяют прийти к предположению, что видовая или сортовая дивергенция осуществляется не только за счёт изменений в ядерных генах, но и за счёт изменений в хлоропластах и митохондриях, причём в одних случаях в большей степени они затрагивают хлоропластные, а в других - митохондриальные гены. Изменение генетического материала этих клеточных структур позволяет косвенно оценить энергоёмкость изучаемых сортов. В связи с этим нами проводилось изучение изменений, происходящих в генетической системе митохондрий и хлоропластов под действием стресса у засухоустойчивого сорта земляники Ленинградская ранняя и слабоустойчивого сорта Боравитская [5].
Таблица 1
Изменение содержания нуклеиновых кислот в проростках земляники в связи с засухой (Мкг % на сухое вещество митохондрий и хлоропластов)
|
№ |
Варианты |
Хлоропластная |
Митохондриальная |
|||||||
|
РНК |
ДНК |
РНК/ ДНК |
РНК |
ДНК |
РНК/ ДНК |
|||||
|
Ленинградская ранная 18 часов стресса |
||||||||||
|
1 |
Контр. |
9154 ±21.3 |
307.6 ±2.7 |
39.8 |
7065 ±73.0 |
177.3 ±6.3 |
39.8 |
|||
|
2 |
ПЭГ |
12107 ±65.4 |
341.9 ±2.5 |
35.4 |
13855 ±35.0 |
367.7 ±5.8 |
37.7 |
|||
|
Боравиткая 18 часов стресса |
||||||||||
|
1 |
Контр. |
16744±15.9 |
328.0±5.0 |
51.0 |
28980±26.5 |
450.7±1.4 |
64.3 |
|||
|
2 |
ПЭГ |
10304±23.7 |
289.6±2.1 |
35.6 |
20240±14.8 |
425.6±2.0 |
47.5 |
|||
Данные приведённые в таблице 1 и на рисунке 1 характеризуют изменения содержания митохондриальной и хлоропластной РНК и ДНК у засухоустойчивого сорта Ленинградская ранняя. Анализ полученных данных показал повышение нуклеиновых кислот в хлоропластах. В процентном отношении к контролю у сорта Ленинградская ранняя содержание РНК возрастает на 86.6%, ДНК на 31.1%. Как видно из полученных данных у засухоустойчивого сорта Ленинградская ранняя наблюдалось и увеличение содержания нуклеиновых кислот в митохондриях.
На фоне засухи у сорта было зафиксировано повышение содержания митохондриального РНК на 136 %, ДНК на 180%.
Изменение содержания нуклеиновых кислот в структурных элементах клетки - митохондриях и хлоропластах, соотношения РНК/ДНК в этих цитоплазматических органеллах указывает на высокий синтез функциональных компонентов. Это позволяет предположить, что у устойчивого сорта Ленинградская ранняя существенный вклад в общий энергетический потенциал клетки вносит и митохондриальная, и хлоропластная системы энергообеспечения.
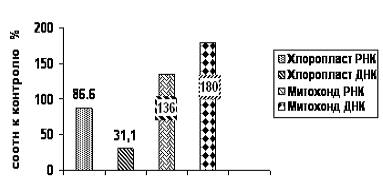
Рис. 1. Диаграмма содержания нуклеиновых кислот в хлоропластах и митохондриях растений земляники сорта Ленинградская ранняя при действии засухи
(по отношению к контролю)
Результаты изучения изменения цитоплазматических генетических систем в условиях засухи проводились также и у слабоустойчивого сорта Боравитская. Как видно из таблицы, у слабоустойчивого сорта наблюдается снижение количества нуклеиновых кислот, как в митохондриях, так и в хлоропластах. Это свидетельствует о том, что у данного образца под действием стресса в хлоропластах и митохондриях идут минимальные биосинтетические процессы. Судя по содержанию РНК и показателям РНК/ДНК, хлоропластая ДНК обладает низкой транскрипционной активностью.
Более наглядную картину уменьшения нуклеиновых кислот в митохондриях и хлоропластах под действием засухи у слабоустойчивого сорта Боравитская представлен на рисунке 2.
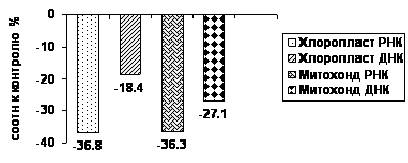
Рис. 2. Диаграмма содержания нуклеиновых кислот в хлоропластах и митохондриях растений земляники сорта Боравитская при действии засухи (по отношению к контролю)
Содержание хлоропластной РНК у сорта Боравитская понижается на 36.8%, ДНК- на 18%. Снижение содержания нуклеиновых кислот у исследуемого сорта, по отношению к контролю в митохондриях составляет в пределах РНК 36.3%, а ДНК - 27.1%. При сравнении с высокоустойчивым сортом у слабоустойчивого биосинтез идёт на более низком уровне и генетическая система митохондрий работает на поддержание уже существующего равновесия.
Анализ изменений содержания нуклеиновых кислот и их функциональной активности в цитоплазматических органеллах в условиях засухи у устойчивого сорта показал, что в наибольшей степени обеспечивает возможность приспособления к неблагоприятному фактору - установления динамического равновесия процессов синтез-распад на новом уровне. Слабоустойчивый сорт характеризуется низкой интенсивностью синтеза нуклеиновых кислот. Полученные нами данные по изменению содержания митохондриальных и хлоропластных нуклеиновых кислот у высокоустойчивого и у слабоустойчивого сортов земляники под действием засухи представляет большой интерес.
Мощность энергетического метаболизма, его потенциальные возможности во многом определяют эффективность биосинтетических процессов, устойчивость и приспособляемость организма к изменяющимся условиям окружающей среды.
СПИСОК ЛИТЕРАТУРЫ
- 1. Алиев Д.А., Казибекова Э.Г. Особенности фотосинтеза высокопродуктивной пшеницы и использование фотосинтетических признаков в селекции // Известие Азерб.,№ 1-6, 2002, с. 20-26
- 2. Алиев Р.Т. Динамика нарушений в метаболизме РНК и ДНК в функции фотосинтетического аппарата под действием стресса у растений ржи / Мат. конф. Экологические аспекты интенс. с/х раст, Пенза, 2002,с.132-136
- 3. Гончарова Э.А, Удовенко Г.В. Онтогенетическая адаптация и регуляция плодоношения при взаимодействии генотип- среда. Доклады Российской Академии с.х наук, 1999, № 6, стр 10-13
- 4. Джавадова Л.Г. и др. Оценка устойчивости пшеницы на ранних этапах её развития / Мат. IV межд. сипм. по нетрадиц. раст., т.1, Пущино, 2001, с.198-201.
- 5. Конарев В.Г., Тютерев С.Л. Методы биохимии и цитохимии нуклеиновых кислот у растений. Ленинград, 1970.
- 6. Кузнецов В.В. и др. Пролин при стрессе: биологическая роль, метаболизм, регуляция - Физиология растений - 1999, т.46, №2, с. 321-336
- 7. Семенов О.Г. Аллоцитоплазматическая пшеницы. В кн.: Биологические основы селекции, Монография. М.: Изд-во РУДН, 2000, с.208.
- 8. Сидоров А.В. Селекция яровой пшеницы на устойчивость к грибным болезням // Селекция и семеноводство, Ж- №3, 2001 с.20-23.
- 9. Шумный В.К. Генная и хромосомная инженерия растений// Сельскохозяйственная биология, №7, 2004, с.12-16.
- 10. Akhundova N.A. Aliev.R.T. Supposed mechanisms of the Humav biofield signal to vegetable cell. FEBS Specif / Meeting cell signaling Mechanisms from Membrane to nucleus, Amsterdam, Nethrlands, 1997, p.96-102.
Библиографическая ссылка
Гаджиева А.Ф ИЗМЕНЕНИЕ ХЛОРОПЛАСТНОЙ И МИТОХОНДРИАЛЬНОЙ ГЕНЕТИЧЕСКОЙ СИСТЕМЫ // Современные проблемы науки и образования. 2009. № 6-3. ;URL: https://science-education.ru/en/article/view?id=1420 (дата обращения: 29.07.2026).