


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
CHARACTERISTIC CALCULATION OF DISCRIMINANT FUNCTION FOR SEPARATION OF ANIMALS ON PHENOTYPIC GROUPS BY THE TIME OF HEXOBARBITAL SLEEP.

Введение
Особенности скорости биотрансформации ксенобиотиков часто являются причиной различного эффекта от действия одной и той же дозы у разных организмов. Скорость биотрансформации определяется активностью цитохрома P-450.
Существуют различные адаптивные механизмы регуляции активности цитохрома P-450. Один из таких механизмов - индукция, то есть повышение концентрации и активности ферментов в ответ на действие ксенобиотика. К механизмам индукции относят: фенобарбиталовый, рифампицин-дексаметазоновый и этаноловый тип [2]. Одним из основных типов является фенобарбиталовый. Даже в небольших концентрациях препараты фенобарбиталового ряда способны к индукции многочисленных изоферментов цитохрома P-450: CYP2B6, CYP2C9, CYP2C18, CYP2C19, CYP3A4, CYP3A5 и др. Таким образом, в случае применения препаратов фенобарбиталового ряда мы можем оценить активность большого числа изоферментов цитохрома Р-450.
Известно, что введение препаратов фенобарбиталового ряда влияет на функции организма по механизму аутоиндукции, что ведет к изменению ответа организма на факторы внешней и внутренней среды. Кроме того, в доступной нам литературе нет единого критерия, позволяющего выделить фенотипические группы животных по активности изоферментов цитохрома P-450 [4].
Следовательно, возникает методическая задача оптимизировать дозировку препаратов фенобарбиталового ряда, применяемых для оценки активности изоферментов цитохрома P-450, с целью снижения эффекта от аутоиндукции и характеристики дискриминантной функции, позволяющей отнести животных к определенной фенотипической группе.
Материалы и методы
Основным прямым методом определения активности цитохрома P-450 у животных является фенотипирование in vivo [1].
В качестве модели оценки активности цитохрома P-450 был использован тест «гексеналового сна». Гексенал вводили крысам внутрибрюшинно в дозе 60 мг/кг. Время сна оценивали с момента потери рефлекса переворачивания до момента его восстановления [5]. Эксперимент проводился на 320 крысах-самцах линии Wistar, полученных из вивария ГБОУ ВПО «ЮУГМУ».
Статистический анализ полученных результатов проводили с помощью пакета программ прикладной статистики SPSS 17. Оценивали параметры распределения, проводили описательную статистику всей совокупности и двухэтапный кластерный анализ [3].
Результаты и обсуждение
На первом этапе мы провели описательную статистику всей совокупности (табл. 1) и проверили совокупность на нормальность распределения (рис. 1).
Таблица 1
Описательная статистика всей совокупности (время «гексеналового сна», мин)
|
N валидных (целиком) |
Среднее |
Стандартная ошибка (Среднее) |
Стандартное отклонение |
Асимметрия |
Стандартная ошибка (Эксцесс) |
Эксцесс |
|
320 |
27,58 |
1,345 |
24,059 |
0,534 |
0,272 |
-0,474 |
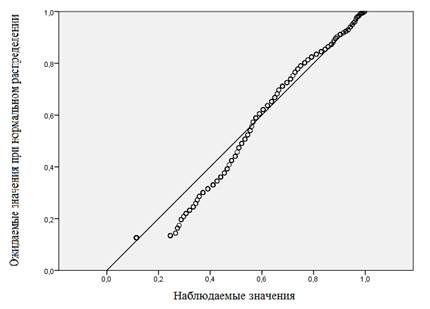
Рис. 1. Вероятностный график распределения всей совокупности (время «гексеналового сна»)
Для определения возможности распределения крыс на фенотипические группы был проведен двухэтапный кластерный анализ с автоматическим определением числа кластеров. Проведенный анализ показал, что действительно по времени «гексеналового сна» крысы распределяются на три фенотипические группы: «быстрые» (ultraextensive metabolism, UM), «медленные» (poor metabolism, PM) и «экстенсивные» (extensive metabolism, EM) метаболизаторы (рис. 2).

Рис. 2. Автоматическое определение количества кластеров
Далее мы провели статистический анализ совокупности, разбитой на кластеры. Результаты анализа для каждого кластера показаны в таблице 2, вероятностные графики для каждого из кластеров приведены на рисунке 3. Из представленных данных следует, что все кластеры характеризуются распределением, близким к нормальному.
Таблица 2
Описательные характеристики для каждого кластера
|
|
«Быстрые» метаболизаторы |
«Медленные» метаболизаторы |
«Экстенсивные» метаболизаторы |
|
|
Среднее |
3,18 |
58,47 |
29,40 |
|
|
95% доверительный интервал для среднего |
Нижняя граница |
2,28 |
55,79 |
27,73 |
|
Верхняя граница |
4,08 |
61,14 |
31,07 |
|
|
Стандартное отклонение |
5,024 |
12,777 |
8,767 |
|
|
Асимметрия |
1,275 |
1,619 |
-0,045 |
|
|
Эксцесс |
0,045 |
2,720 |
-1,356 |
|
|
Стандартная ошибка |
0,455 |
1,347 |
0,844 |
|
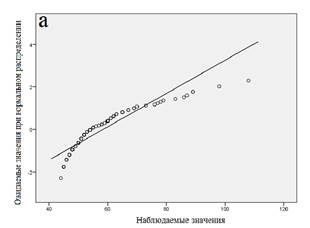
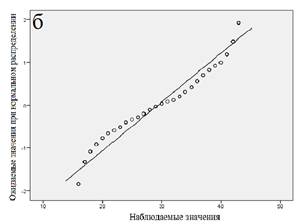
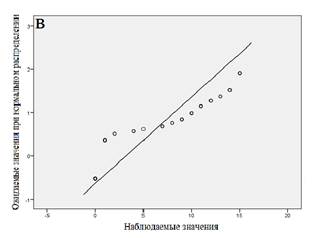
Рис. 3. Вероятностные графики (а - «медленные» метаболизаторы; б - «быстрые» метаболизаторы; в - «экстенсивные» метаболизаторы)
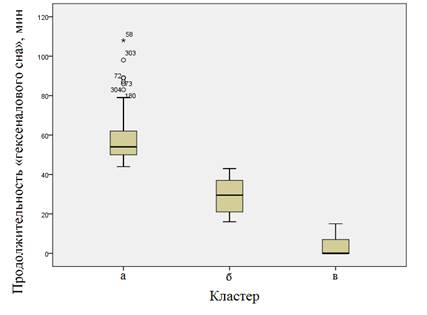
Рис. 4. Диаграмма распределения продолжительности «гексеналового сна» в зависимости от кластерной принадлежности (а - «медленные» метаболизаторы; б - «экстенсивные» метаболизаторы; в - «быстрые» метаболизаторы)
На рисунке 4 показаны диаграммы распределения продолжительности сна в зависимости от кластерной принадлежности.
На втором этапе статистической обработки результатов с целью расчета вероятности ошибочного распределение крыс на фенотипические группы по скорости «гексеналового сна» был проведен дискриминантный анализ. Дискриминантный анализ выполнялся для трех кластеров с расчетом нормированного и ненормированного коэффициента канонической дискриминирующей функции. Также рассчитывались значения центров групповых центроидов, которые позволяют определить принадлежность к кластеру. Поскольку переменная только одна - время «гексеналового сна», то и дискриминантная функция также была одна. На рисунке 5 приведены значения функции в центроидах групп для трех кластеров, позволяющие рассчитать принадлежность конкретного животного к тому или иному кластеру.
|
«быстрые» метаболизаторы |
«экстенсивные» метаболизаторы |
«медленные» метаболизаторы |
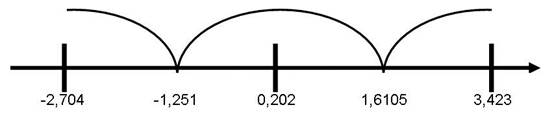
│- значения дискриминантной функции в центроидах кластера.
![]()
- интервал значений числовой прямой.
Рис. 5. Расположение по числовой прямой значений в центроидах дискриминантной функции (время «гексеналового сна») каждого из кластеров.
Как видно из рисунка 5, значения дискриминантной функции более 1,6105 позволяют отнести опытное животное к кластеру «медленных» метаболизаторов, значения дискриминантной функции от -1,251 до 1,6105 – к кластеру экстенсивных метаболизаторов и менее -1,251 – к кластеру «быстрых» метаболизаторов.
В таблице 3 приведены коэффициенты канонической дискриминантной функции.
Таблица 3
Коэффициенты канонической дискриминантной функции времени «гексеналового» сна
|
|
Значение |
|
Время гексеналового сна (k) |
0,111 |
|
Константа дискриминантной функции (С0) |
-3,056 |
Дискриминантная функция рассчитывается следующим образом: F(t)=C0+kt. Например, пусть время сна составило 10 минут, тогда F(t)= -3,056 +0,111• t, где t – время «гексеналового сна» в минутах, следовательно F(t)= -3,056 +0,111•10=-1,946. Соотносим данное значение с рисунком 5 и делаем вывод, что данное экспериментальное животное относится к группе «быстрых» метаболизаторов.
Таблица 4
Результаты дискриминантного анализа
|
|
Кластер |
Центроиды
|
Предсказание принадлежности к группе (%) |
Итого (%) |
||
|
«медленные» метаболизаторы |
«экстенсивные» метаболизаторы |
«быстрые» метаболизаторы |
||||
|
Исходные |
«медленные» метаболизаторы |
3,423 |
98,9 |
1,1 |
0,0 |
100,0 |
|
«экстенсивные» метаболизаторы |
0,202 |
0,0 |
94,4 |
5,6 |
100,0 |
|
|
«быстрые» метаболизаторы |
-2,704 |
0,0 |
100,0 |
100,0 |
100,0 |
|
Результат дискриминантного анализа свидетельствует о том, что в общем случае 97,8% исходных наблюдений классифицировано правильно.
Таким образом, все животные по времени «гексеналового сна» были распределены по трем фенотипическим группам, что соотносится с данными литературы, и для каждой для них определены границы.
В результате проведенных исследований была рассчитана и охарактеризована дискриминантная функция, что позволило утверждать, что при использовании гексенала в концентрации 60 мг/кг сохраняется разделение животных на три фенотипические группы по активности изоферментов цитохрома P-450.
Рецензенты:
Головнева Е.С., д.м.н., профессор кафедры нормальной физиологии ГБОУ ВПО «ЮУГМУ» Минздрава России, г. Челябинск.
Рябинин В.Е., д.м.н., профессор кафедры биохимии ГБОУ ВПО «ЮУГМУ» Минздрава России, г. Челябинск.
Библиографическая ссылка
Козочкин Д.А., Болотов А.А., Цейликман В.Э., Тишевская Н.В., Абрамовских О.С., Никитина А.А., Комелькова М.В., Мишарина М.Е. РАСЧЕТ ХАРАКТЕРИСТИК ДИСКРИМИНАНТНОЙ ФУНКЦИИ ДЛЯ РАЗДЕЛЕНИЯ ЖИВОТНЫХ НА ФЕНОТИПИЧЕСКИЕ ГРУППЫ ПО ВРЕМЕНИ ГЕКСЕНАЛОВОГО СНА. // Современные проблемы науки и образования. 2014. № 1. ;URL: https://science-education.ru/en/article/view?id=11945 (дата обращения: 27.06.2026).