


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
MORPHOLOGICAL CHARACTERISTIC ТРИХОМ OF KINDS OF SORT JUGLANS GROWING IN THE CONDITIONS OF THE SOUTH OF CENTRAL RUSSIAN UPLAND

Введение
У растений из сем. Juglandaceae поверхность листьев и перикарпия покрыта железистыми и нежелезистыми трихомами нескольких морфологических типов. Одни авторы выделяют 4 основных типа [2], другие – 6, при этом учитывается внутривидовая, сезонная, географическая и экологическая изменчивость, распределение на адаксиальной и абаксиальной поверхностях листовой пластинки [8]. Несмотря на то, что существует единая база данных о трихомах и генах вовлеченных в синтез метаболитов трихом [6], в работах разных авторов основные типы трихом имеют разные обозначения. Важное значение имеет идентификация трихом в систематическом отношении [6], в том числе для определения ископаемых видов [7,10]. Трихомы играют важное значение в поддержании водного баланса и в регулировании температуры листа [4,12], так же они выполняют защитную роль по отношению к возбудителям болезней, способствуют опылению в фазу цветения, оказывают влияние на фотосинтез [12].
Цель настоящего исследования – идентифицировать трихомы поверхности листа интродуцированных в Белгородскую область видов рода Juglans, обобщить и дополнить существующие классификации.
Материалы и методы. Объектами исследования стали растения семи видов рода Juglans, произрастающих в Ботаническом саду БелГУ, 2001 года посадки: J. regia L., J. manshurica Max., J. siboldiana Maxim., J. cjhdiformis Max., J. nigra L., J. cinerea L., J. rupestris Engelm. Контроль – растения вида J. regia L., произрастающий в течение длительного времени в Белгородской области.
Учитывалось происхождение видов. Для исследования отбирались листья с годичных приростов (7-й от основания прироста) с учетом их морфологического адреса, освещенности в утренние часы. Консервация листьев проводилась общепринятым способом в смеси: спирт, вода, глицерин (1:1:1) [3]. Приготовление препаратов эпидермиса проводилось по модифицированной методике [3].
Изучение эпидермиса и его структур проводилось с помощью СМ «Биолам С 13», Микромед-5, бинокуляра МБС 10 и РЭМ Quanta 200 3D в центре коллективного пользования научным оборудованием БелГУ.
При составлении классификации учитывались признаки зрелых, полностью сформированных структур. Учитывали форму трихом, число клеток, измеряли морфологические параметры, распределение на адаксиальном и абаксиальном эпидермисе листовой пластинки. Рассчитывали площадь пельтатных трихом и коэффициент их эксцентричности [4].
Результаты и обсуждение
У представителей семейства Juglandaceae вегетативные и репродуктивные органы покрыты морфологически разнообразными трихомами [2]. А.Tachtadjan характеризует растения семейства Juglandaceae наличием на поверхности листьев погруженных пельтатных трихом и нежелезистых волосков [11. David L. Dilcher (1986), использует особенности листовой поверхности для идентификации растений в палеоботанике [7]; автор описывает типы трихом известных растений семейства Juglandacea, он выделяет три типа трихом: простые одноклеточные («simple»), большие и маленькие пельтатные трихомы («large» and «small pellate»). Пельтатные трихомы характерны для растений семейства Juglandaceae [15]. David L. Dilcher отмечает их у растений трех родов Alforoa, Oreomunnea, Engelhardia [13]. Растения родов Alforoa, Oreomunnea могут отличаться наличием или отсутствием простых одноклеточных трихом [10].
Для описания разнообразия и морфофункциональных особенностей трихом в роде Salvia Е.В. Байкова [1] использует маркировку, которую мы применяем в нашем исследовании - пояснения: Г1 – ножка 2-хклеточная, головка 2-4-хклеточная;Г2 – ножка 5-клеточная, головка 8-9-клеточная;Г3 – ножка 8-9-клеточная, головка 4-6-клеточная.
В настоящем исследовании проводится классификация типов трихом рода Juglans, учитываются известные варианты классификаций, дополняются существующие сведения, дается характеристика различным подтипам (морфологические параметры, распределение) (табл. 2). Учитываются такие характеристики как число клеток в структурах трихом, разветвленность.
Таблица 2 - Трихомы абаксиальной поверхности листа видов рода Juglans
|
|
Нежелезистые |
Железистые |
||||||||||
|
Однорядные |
Пучковые |
Головчатые |
Пельтатные |
|||||||||
|
Простые
КП1 |
Сложн-ые КС1 |
Сложн-ые ПУ2 |
Много-лучевые ПУ1 |
Малень-кие Г1 |
Большие |
маленькие |
большие |
|||||
|
Г2 |
Г3 |
4-кл. Ж1 |
6-8-кл. Ж2 |
Много-кл.Ж3 |
||||||||
|
Абаксиальный эпидермис листа |
||||||||||||
|
J. cinerea |
+ |
|
+ |
+ |
+ |
+ |
+ |
+ |
|
+ |
||
|
J. siboldiana |
+ |
|
+ |
+ |
+ |
+ |
|
+ |
+ |
+ |
||
|
J. manshurica |
+ |
+ |
+ |
|
|
+ |
|
+ |
|
+ |
||
|
J. rupestris |
|
+ |
|
|
|
+ |
+ |
+ |
|
+ |
||
|
J. cjhdiformis |
|
+ |
+ |
|
+ |
+ |
+ |
|
|
+ |
||
|
J. regia |
|
|
|
|
|
|
|
|
|
+ |
||
|
J. nigra |
+ |
|
|
|
|
+ |
|
+ |
|
+ |
||
|
|
Адаксиальный эпидермис листа |
|||||||||||
|
J. cinerea |
+ |
|
+ |
|
+ |
+ |
+ |
+ |
|
+ |
||
|
J. siboldiana |
+ |
|
+ |
+ |
|
+ |
+ |
+ |
|
+ |
||
|
J. manshurica |
+ |
|
+ |
|
|
|
|
+ |
|
+ |
||
|
J. rupestris |
|
|
|
|
|
|
|
+ |
|
+ |
||
|
J. cjhdiformis |
|
+ |
+ |
|
|
|
+ |
|
|
+ |
||
|
J. regia |
|
|
|
|
|
|
|
|
|
+ |
||
|
J. nigra |
|
|
|
|
|
+ |
|
+ |
|
+ |
||
Нежелезистые одноклеточные трихомы (рис. 2), которые похожи по описанию на «игольчатые» [16], обнаружены на адаксиальном и абаксиальном эпидермисе ореха серого, серповидно изогнуты (рис. 1, КП1) (длина 319,23±44,13мкм). У ореха Зибольда кроющие одноклеточные трихомы располагаются на абаксиальном и адаксиальном эпидермисе редко, только на жилках (длина 327,39±25,84 мкм). У ореха маньчжурского располагаются равномерно на абаксиальном и на адаксиальном эпидермисе (длина 373,88±44,32 мкм). На абаксиальном эпидермисе ореха черного редко встречаются одноклеточные кроющие трихомы конической формы, длина 91,19±10,11 мкм.
|
|
|
|
КП1. Серповидно изогнутые, адаксиальный эпидермис (×500) J. cjhdiformis |
КП2 Прямые абаксиальный эпидермис (×1200) J. rupestris |
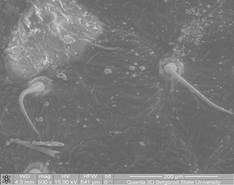

Рис.1. Форма нежелезистых одноклеточных трихом растений рода Juglands
Сложные однорядные неветвистые конические волоски у основания изогнутые обнаружены на адаксиальном и абаксиальном эпидермисе ореха сердцевидного (рис. 2, КП), длина от 122 до 200 мкм. На адаксиальном эпидермисе они распределены равномерно. На абаксиальном эпидермисе их концентрация больше на жилках. У ореха скального многоклеточные однорядные трихомы обнаружены только на абаксиальном эпидермисе. Они прямые, распределены равномерно, длина 150,56±15,5 мкм.
|
Гр. |
Тип |
Подтип |
||
|
Нежелезистые |
Неветвистые волоски |
|
|
|
|
Простые (одноклеточные) конические изогнутые КП1 (×2240) |
Простые (одноклеточные) конические прямые КП2 (×1120) |
Сложные однорядные неветвистые конические КС1 (×800) |
||
|
Пучковатые |
|
|
|
|
|
Многолучевые ПУ1 (×800) |
Сложные пучковатые ПУ2 (×1000) |
Сложные пучковатые ПУ2 (×1600) |
||
|
Железистые |
Головчатые неветвистые волоски |
|
|
|
|
Ножка из 2 конических клеток, головка 2-4 клетки Г1 (×2100) |
Ножка из 5 конических клеток. Головка 8-9-клеточная Г2 (×2100) |
Ножка из 8-9 конических клеток. Головка 4-6-клеточная Г3 (×1000) |
||
|
Пельтатные железки |
|
|
|
|
|
4-х клеточная Ж1 (×2000) |
6-8-клеточная Ж2 (×2000) |
Многоклеточная Ж3 (×1600) |
||
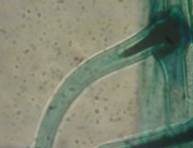

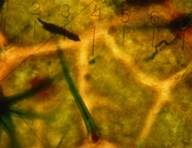


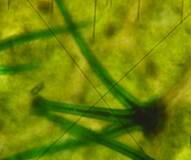


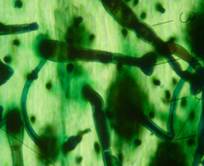
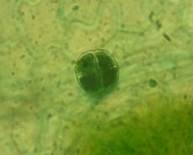

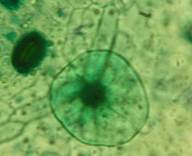
Рис. 2. Морфологическая классификация трихом рода Juglands
Нежелезистые одиночные трихомы («Solitary»), длиной 450-1600 мкм, описанные James Hardin (1984) [8], не были обнаружены.
Нежелезистые сложные пучковатые трихомы и многолучевые имеют вид нескольких одноклеточных и многоклеточных однорядных нежелезистых трихом, имеющих общее основание (рис. ПУ 2). Длина ответвлений варьирует от 125 до 560 мкм. Диаметр многоклеточного основания 47,24±2,50 мкм. У сложных пучковых трихом число ответвлений 2-8 шт., у многолучевых – 8-15 шт.
Пучковые трихомы обнаружены на абаксиальном эпидермисе ореха серого, Зибольда, маньчжурского, сердцевидного. Многолучевы трихомы обнаружены на жилках на адаксиальной и абаксиальной поверхностях ореха Зибольда.
Длина самого большого ответвления пучковой трихомы у ореха серого 553,04±143,11 мкм (число ответвлений 5,3±0,503), у ореха Зибольда – 302,48±76,83 мкм (число ответвлений 7,10±1,13), у ореха манчьжурского - 498,44±129,45 мкм (число ответвлений 3,40±0,340). Обнаружена высокозначимая линейная корреляция между длиной ответвлений и их числом (r=0,833).
James Hardin отметил на поверхности эпидермиса листьев растений Carya tomentosa приподнятые пучковые трихомы, которые находятся на многоклеточном основании [15]. Для всех видов нами отмечены только сидячие пучковые трихомы.
Основные клетки эпидермиса, прилегающие к основанию трихом, обнаруживают иную форму и размеры, чем остальные клетки эпидермиса. Они имеют трапециевидную форму. Своими малыми основаниями они примыкают к основанию волоска, создавая вид радиального расположения антиклинальных оболочек клеток, образующих ребра трапеции. Клетки у основания волоска, покрыты кутикулой, внешне напоминая «валик», диаметр валика – 50,05±10,5 мкм.
Головчатые трихомы обнаружены на адаксиальном и абаксиальном эпидермисе всех видов ореха, кроме грецкого.
Головчатые трихомы на короткой ножке (рис. 2, Г1). Орех серый имеет на адаксиальной и абаксиальной поверхности листа маленькие головчатые трихом, ножка двуклеточная, головка имеет от 2-х до 4-х клеток. Их концентрация больше на жилках.
На абаксиальном эпидермисе ореха Зибольда на жилках обнаружены головчатые трихомы на двуклеточной ножке. Головки крупные, в них от 6 до 12 клеток, большая их часть повреждена. Орех сердцевидный имеет трихомы с двухклеточной ножке, 3-4-х клеточной головкой на абаксиальном эпидермисе.
Головчатые трихомы на длинной ножке из 5 клеток (длина 120,42±15,70 мкм), 3-4 клеточной головкой (диаметр 93,1±3,17 мкм) расположены на абаксиальном и адаксиальном эпидермисе ореха серого (рис. 2, Г2). На абаксиальном эпидермисе они обнаруживаются только на жилках. У ореха Зибольда такие железистые трихомы обнаружены на абаксиальном эпидермисе на жилках (длина ножки 54,37±3,14 мкм, диаметр головки 77,10±5,58 мкм). Орех черный имеет головчатые трихомы на ножке из 6 клеток (длина 115,17±20,15 мкм), головка из 7-8 клеток (диаметр 57,48±5,87 мкм) на адаксиальном эпидермисе (только на жилках) и на абаксиальном эпидермисе (равномерно расположены) (рис. 2, Г3). Орех скальный имеет данный подтип железистых трихом на абаксиальном и адаксиальном эпидермисе. Число клеток в ножке варьирует от 3 до 7 (длина 56,35±8,24 мкм), большая часть головок разрушена. Орех манчьжурский имеет головчатые трихомы на длинной ножке на адаксиальном и абаксиальном эпидермисе, расположенные только на жилках. Длина ножки 44,69±5,45 мкм, диаметр головки 64,49±11,11 мкм. На адаксиальном и абаксиальном эпидермисе ореха сердцевидного головчатые трихомы имеют в ножке 3-5 клеток.
Пельтатные трихомы характерны для растений семейства Juglandacea. В литературе название этого типа трихом имеет ряд синонимов the «peltate glands», «resin dots», «scales» и «lepidote scales». Они встречаются с разной частотой у всех изученных нами видов. Отмечены 4-х, 8-клеточные (маленькие), а также многоклеточные (большие) пельтатные трихомы. У ореха серого четырехклеточные пельтатные трихомы отмечены на адаксиальном эпидермисе редко, на абаксиальном эпидермисе – равномерно. У ореха Зибольда мелкие пельтатные железки на адаксиальном эпидермисе редко встречаются, а на абаксиальном – четырех- и восьмиклеточные (S 509,17±38,54 мкм2) (табл. 3). У ореха черного мелкие пельтатные трихомы обнаружены на абаксиальном и адаксиальном эпидермисе, диаметр 400,74±39,20 мкм. У ореха манчьжурского мелкие пельтатные трихомы располагаются на адаксиальном и абаксиальном эпидермисе редко. Многоклеточные пельтатные трихомы отмечены у всех изучаемых видов рода Juglans. У ореха грецкого отмечен только этот подтип трихом, 2,96±0,249 штук на 1 мм2 абаксиального эпидермиса. Наибольшая площадь многоклеточных пельтатных трихом отмечена у ореха манчьжурского, Зибольда, а наименьшая у ореха сердцевидного, грецкого и скального. Наиболее округлую форму имеют пельтатные трихомы ореха Зибольда и грецого, что соответствует большей их наполненности секретом.
Таблица 3 - Характеристика многоклеточных пельтатных трихом растений рода Juglans
|
Вид |
Площадь пельтатных железок, ×103 мкм2 |
Площадь субкутикулярной полости пельтатных железок, мкм2 |
Коэффициент эксцентричности пельтатной железки |
Коэффициент эксцентричности субкутикулярной полости железки |
|
J. cinerea |
9,845±0,321 * |
5,5095±0,244 ** |
0,495±0,027* |
0,505±0,032** |
|
J. siboldiana |
10,045±0,203 ** |
4,259±0,290 ** |
0,325±0,030 |
0,420±0,035 |
|
J. manshurica |
11,876±0,131 ** |
3,592±0,129 ** |
0,424±0,017 |
0,227±0,016 |
|
J. rupestris |
3,363±0,410 ** |
1,112±0,133 * |
0,560±0,039** |
0,558±0,046 |
|
J. cjhdiformis |
0,758±0,056 ** |
0,339±0,010 ** |
0,508±0,029* |
0,500±0,027 |
|
J. regia |
1,858±1,338 |
0,162±0,117 |
0,389±0,032 |
0,494±0,047 |
На поверхности пельтатных трихом кутикулярный слой более толстый, он формирует особый рельеф. Чаще всего многоклеточные пельтатные трихомы имеют радиальные складки вдоль антиклинальных стенок (орех серый, сердцевидный). У ореха черного кутикула более тонкая, восковой налет на поверхности эпидермы не выражен. Кутикула в большей степени спадает вокруг субкутикулярной полости (рис. 4 Б).
|
|
|
|
А. J. cinerea (×2000) |
Б. J. nigra (×2000) |


Рис. 4. Рельеф поверхности кутикулярного слоя пельтатных трихом абаксиальной поверхности листа растений рода Juglands
По-видимому, складчатость кутикулы является следствием сжатия имевшейся в головке субкутикулярной полости. Особенности поверхностного рельефа обусловлены не только локализацией и размером субкутикулярной полости, но также толщиной кутикулы. Очевидно, что более толстая кутикула препятствует образованию складок [2].
В отсутствии внешних воздействий поверхность пельтатных трихом остается целостной, в противном случае приподнятая кутикула полностью отрывается и оторванный секреторный продукт оказывается снаружи [13]. Существует мнение [14] о том, что факторами, вызывающими разрыв кутикулы, является высокая температура, низкая влажность воздуха и механическое давление. Такой же механизм секреции, сопровождающийся отрывом кутикулы, наблюдается в пельтатных железках листьев R. americanum [8], Ocimum basilicum [9] и в трихомах завязи Zeheria mantana [11].
Функции трихом, расположенных на поверхности околоплодников и способных синтезировать вторичные метаболиты, многообразны. В период цветения эфирные масла, входящие в состав секрета, капли слизи, а также яркое окрашивание привлекают насекомых-опылителей. Фенольные и терпеноидные соединения защищают вегетативные и генеративные органы от фитопатогенов, как это было установлено для растений из других систематических групп [11]. Трихомы играют определенную роль как в механической защите плодов от травоядных насекомых, так и в предотвращении иссушения их поверхности [3].
Заключение
Листья растений видов J. regia, J. manshurica, J. siboldiana, J. cjhdiformis, J. nigra, J. cinerea, J. rupestris опушены. Трихомы, расположенные на абаксиальной и адаксиальной поверхности листа классифицируют на две группы: железистые и нежелезистые. К нежелезистым трихомам относятся простые (одноклеточные) конические трихомы, которые по форме делятся на прямые и серповидно изогнутые. Также среди нежелезистых трихом впервые описаны многоклеточные однорядные конические трихомы. К этой же группе трихом относятся пучковые сложные с 5-7-ю ответвлениями и многолучевые с 8-15 ответвлениями (обнаружены только на листьях J. siboldiana и на абаксиальной поверхности листа J. cinerea). К группе железистых трихом относятся головчатые трихомы: маленькие (с двухклеточной ножкой, 2-4-хклеточной головкой), большие (с 5-клеточной ножкой, 8-9-клеточной головкой и трихомы с 8-9-клеточной ножкой и 4-6-клеточной головкой). К железистым трихомам относятся пельтатные железки, среди которых выделены маленькие 4-х клеточные железки, 6-8-клеточные железки и большие многоклеточные железки.
Поверхность волосков не образует складок, однако на ней хорошо выявляется микрорельеф, спирально ориентированный по отношению к вертикальной оси клеток. Многоклеточные пельтатные трихомы морфологически отличаются. На поверхности эпидермиса листов J. nigra с недостаточно выраженным восковым налетом кутикула «спадает», субкутикулярная полость хорошо выражена. Особенности поверхностного рельефа обусловлены не только локализацией и размером субкутикулярной полости, но и также толщиной кутикулы.
Исследования выполнены в рамках реализации государственного задания Министерства образования и науки РФ Белгородским государственным национальным исследовательским университетом на 2013 год (№ проекта 5.1739.2011).
Рецензенты:
Ткаченко И.К., д.с.-х.н., профессор, профессор кафедры анатомии и физиологии живых организмов Биолого-химического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород.
Лазарев А.В., д.б.н., доцент, профессор кафедры биотехнологии и микробиологии Биолого-химического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород.
Библиографическая ссылка
Сорокопудов В.Н., Назарова Н.В., Кузнецова Т.А., Колесников Д.А. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ТРИХОМ ВИДОВ РОДА JUGLANS, ПРОИЗРАСТАЮЩИХ В УСЛОВИЯХ ЮГА СРЕДНЕРУССКОЙ ВОЗВЫШЕННОСТИ // Современные проблемы науки и образования. 2013. № 5. ;URL: https://science-education.ru/en/article/view?id=10466 (дата обращения: 07.07.2026).