



Системы токсин-антитоксин были первоначально обнаружены на бактериальных плазмидах как генетические элементы, обеспечивающие стабильное наследование плазмид в бактериальной популяции [4]. Как правило, эти системы представляют собой оперон из двух генов, первый из которых кодирует антитоксин, а второй - токсин [3]. Стабильная передача плазмиды в популяции обеспечивается за счет гибели клеток, в которых произошла случайная потеря плазмиды (post-segregational killing). Механизм специфической гибели бесплазмидных клеток определяется различием уровней стабильности токсина и антитоксина, - токсин является стабильным белков, а антитоксин быстро деградирует под действием внутриклеточных протеаз. Клетки, в которых произошла потеря плазмиды при делении, не будут содержать токсин-антитоксинового оперона и после быстрой инактивации антитоксина будут убиты оставшимся в клетке токсином [3].
По мере расшифровки полных геномов бактерий и архей на протяжении последних 15 лет стало ясно, что большое число разнообразных токсин-антитоксиновых систем кодируется не только плазмидами, но и хромосомами. Функции этих хромосомных локусов не очевидны и являются предметом обсуждения [10]. Высказываются предположения, что эти системы могут являться функциональным аналогом апоптоза эукариотических клеток и/или обеспечивать ответ бактериальной клетки на стрессовые условия, при которых происходит активация протеаз и деградация антитоксина. В этом случае клетка может переводиться в «спящее» состояние, в котором она будет находиться до прекращения действия стрессовых факторов. Обеспечивающие такую «стрессоустойчивость» токсин-антитоксиновые системы могут являться важными факторами патогенности, например, геном возбудителя туберкулеза Mycobacterium tuberculosis кодирует более 60 токсин-антитоксиновых систем, а в геноме непатогенной микобактерии Mycobacterium smegmatis имеется лишь одна такая система [7].
По своему составу и механизму действия токсин-антитоксиновые системы могут быть разделены на три группы [10]. В системах типа I экспрессия гена токсина ингибируется антисмысловой РНК, транксрибируемой в противоположном направлении, т.е. антитоксином является молекула РНК. Чаще встречаются системы типа II, в которых токсин и антитоксин являются белками, их взаимодействие приводит к формированию комплекса, не проявляющего токсической активности. Наконец, в системах типа III антитоксин является молекулой РНК, которая взаимодействует с белком-токсином и ингибирует его активность.
Как было отмечено выше, генетические модули токсин-антитоксин встречаются в большинстве просеквенированных геномов бактерий и архей, причем их функции могут быть связаны с регуляцией ответа клетки на стрессовые условия. Недавно мы определили нуклеотидную последовательность генома термоалкалофильной бактерии Thermosyntropha lipolytica [1]. Это относящийся к порядку Clostridiales микроорганизм [9] растет в экстремальных условиях высоких температур (от 52 до 70 оС) при щелочных значениях pH (7.15 - 9.5). Естественная вариация параметров температуры и рН в содовом озере приводит к периодическому попаданию бактерии в стрессовые условия, в которых функции токсин-антитоксиновой системы могут быть ей полезными.
Цель исследования: идентифицировать и охарактеризовать биоинформационными методами токсин-антитоксиновую систему, кодируемую геномом T. lipolytica.
Материалы и методы исследования
Штамм T. lipolytica был выделен В. А. Светличным из щелочного озера Богория в Кении [9] и предоставлен нам для исследований. Нуклеотидная последовательность генома этой бактерии была определена нами методом пиросеквенирования на геномном анализаторе GS FLX [1]. Поиск открытых рамок считывания, способных кодировать белки, осуществляли с помощью программы Genemark. Для предсказания функций белков соответствующие аминокислотные последовательности сравнивали с базой данных NCBI с помощью BLASTP. Анализ вторичной структуры белков проводили с помощью PSIPRED [5].
Результаты и их обсуждение
Поиск гомологов белковых продуктов, предсказанных открытых рамок считывания, кодируемых в геномных последовательностях T. lipolytica по базе данных NCBI позволил идентифицировать локус, содержащий гомологи генов бактериальных токсина и антитоксина. Эти гены, toxA (антитоксин) и toxB (токсин), образуют оперон, структура которого характерна для токсин-антитоксиновых систем (рис. 1). Эти гены перекрываются на 11 нуклеотидов (включая стоп-кодон первого гена), а в 5'-концевой области оперона располагается вероятный промотор. Интересной особенностью является наличие вблизи района (-10) промотора инвертированного повтора TTAGCATAA. Вероятно, эта последовательность является сайтом связывания комплекса токсин-антитоксин, таким образом репрессирующего промотор оперона. Такой механизм регуляции по принципу отрицательной обратной связи может поддерживать постоянную концентрацию токсина и антитоксина в клетке, устраняя их случайные флуктуации.
Гомологичные на уровне нуклеотидных последовательностей опероны были обнаружены нами в геномах, родственных T. lipolytica бактерий, - Thermoanaerobacterium thermosaccharolyticum DSM 571, Thermoanaerobacterium saccharolyticum JW/SL-YS485, Pelotomaculum thermopropionicum SI, Syntrophothermus lipocalidus DSM 12680 и Thermincola sp. JR. Высокий уровень гомологии наблюдался не только в пределах кодирующих последовательностей toxAB генов, но и в промоторной области. В частности, последовательность TTAGCATAA сохранялась в неизменном виде у всех перечисленных бактерий, несмотря на более низкий уровень гомологии участков, расположенных между этим сайтом и стартовым кодоном toxA (рис. 1), что свидетельствует о функциональной значимости этого сайта.
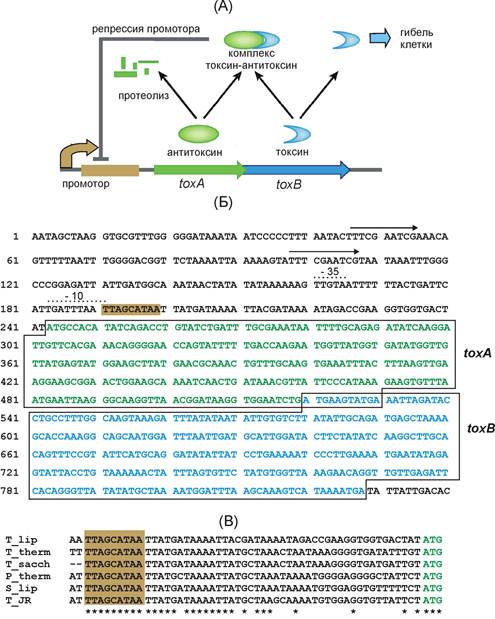
Рис. 1. (А) Структура и схема функционирования toxAB оперона T. lipolytica. (Б) Нуклеотидная последовательность toxAB оперона T. lipolytica. Районы -35 и -10 промотора показаны пунктирными линиями, стрелками показана повторяющаяся последовательность с неизвестной функцией. (С) Сравнение нуклеотидных последовательностей промоторных областей токсин-антитоксиновых оперонов T. lipolytica (T_lip), - T. thermosaccharolyticum (T_therm), T. saccharolyticum (T_sacch), P. thermopropionicum (P_therm), S. lipocalidus (S_lip) и Thermincola sp. JR (T_JR).
Ген toxB кодирует небольшой белок, состоящий из 102 аминокислотных остатков и имеющий молекулярную массу 12 кД. Это щелочной белок (pI 9.13), что характерно для токсинов в токсин-антитоксиновых системах [10]. Напротив, антитоксины, как правило, характеризуются низким pI, что обеспечивает стабилизацию комплекса токсин-антитоксин. Поиск гомологов ToxB в базах данных показал, что он относится к распространенному у бактерий семейству RelE/ParE токсинов (COG3668). Помимо гомологии на уровне аминокислотных последовательностей, анализ вторичной структуры ToxB выявил наличие тех же элементов (альфа-спирали и бета-слои), характерных для прототипа семейства - белка ParE плазмиды RK2 (рис. 2). Несмотря на сходство аминокислотных последовательностей токсинов семейства RelE/ParE, их мишени в клетки могут быть различными. Так, ParE плазмиды RK2 ингибирует ДНК-гиразу и, таким образом, блокирует репликацию хромосомы, а RelE Escherichia coli связывается с рибосомой и ингибирует трансляцию, активируя разрезание находящейся на рибосоме РНК [8]. Определение клеточной мишени токсина ToxB возможно лишь экспериментальным путем.

Рис.2. Выравнивание аминокислотных последовательностей ToxA и ParE белка плазмиды RK2. Вторичная структура ToxA предсказана с помощью PSIPRED [5]
Первый ген оперона, toxA, кодирует белок из 95 аминокислотных остатков с молекулярной массой 11,1 кД и pI 5.53. Поиск гомологов ToxА в базах данных показал, что он относится к семейству YefM, прототип которого был обнаружен у E. coli. К этому же семейству относится антитоксин Phd - компонент системы phd-doc бактериофага Р1, обеспечивающей стабильное наследование его профага [4]. Антитоксины группы YefM гомологичны антитоксинам RelB токсин-антитоксиновых систем ReB-RelE и поэтому часто относятся к этому семейству [3]. Характерной особенностью YefM антитоксина E. coli, а также Phd, является редкий тип конформации белка - в нативном состоянии он является «развернутой» (unfolded) молекулой, чувствительной к действию протеаз [2]. Комплекс токсина с антитоксином, напротив, имеет компактную конформацию и отличается высокой стабильностью. Однако среди YefM антитоксинов имеются и структурированные белки, например, YefM антитоксин из M. tuberculosis [10].
Выводы
Геном термоалкалофильной бактерии T. lipolytica содержит оперон toxAB, кодирующий токсин-антитоксиновую систему ReB-RelE типа. Функции этого оперона могут быть связаны с регуляцией физиологического состояния клетки в экстремальных условиях при изменении показателей температуры и рН воды содового озера.
Работа была выполнена при выполнена при финансовой поддержке Министерства образования и науки РФ (ФЦП «Научные и научно-педагогические кадры инновационной России» на 2009 - 2013 годы, государственный контракт П479).
Рецензенты:
- Бонч-Осмоловская Елизавета Александровна, д.б.н., зам. директора по научной работе ФГБУН Института микробиологии им. С. Н. Виноградского Российской академии наук, г. Москва.
- Игнатов Александр Николаевич, д.б.н., ведущий научный сотрудник ФГБУН Центра «Биоинженерия» Российской академии наук, г. Москва.
Библиографическая ссылка
Марданов А.В., Равин Н.В. ИДЕНТИФИКАЦИЯ ТОКСИН-АНТИТОКСИНОВОГО ОПЕРОНА В ГЕНОМЕ ТЕРМОАЛКАЛОФИЛЬНОЙ БАКТЕРИИ THERMOSYNTROPHA LIPOLYTICA // Современные проблемы науки и образования. 2012. № 5. ;URL: https://science-education.ru/ru/article/view?id=7165 (дата обращения: 01.07.2026).