



Изучение фотосинтеза чая требует проведения тщательных систематических исследований хода этого процесса в зависимости от разнообразных внешних факторов, влияющих на жизнедеятельность чайного растения. Применительно к культуре чая наиболее полные исследования процесса фотосинтеза в условиях Западной Грузии были проведены профессорами М.М. Гочолашвили и Ш.Г. Залдастанишвили [7]. Авторам удалось убедительно доказать, что азотные, фосфорные и калийные удобрения значительно повышают интенсивность фотосинтеза и дыхания листьев чая, произрастающего в типичных условиях влажных субтропиков. Следовательно, приступая к изучению фотосинтеза растений чая в наших условиях, мы имели отправную информацию об этом процессе, что и позволило избежать некоторых методических ошибок.
Материалы и методы исследования
Поставленные задачи решались на уровне полевых и лабораторных исследований с использованием методов математического моделирования и статистики. Опыт по некорневой подкормке микроэлементами заложен в 1996 году на участке клоноиспытания АОЗТ «Дагомысчай», п. Уч-Дере, площадью 0,4 га на растениях чая сорта «Каратум», 1990 года посадки. Интенсивность фотосинтеза измеряли в неотделенных от растений листьях в токе естественного воздуха с помощью кондуктометрического прибора В.Л. Вознесенского [6], усовершенствованного и изготовленного снс лаборатории физиологии и биохимии растений Пилипенко В.Г. Усовершенствование затронуло изменение конструкции листовой камеры, в связи с тем, что средняя площадь листа чая лежит в пределах 30 - 40 см2 и интенсивность фотосинтеза чая по литературным данным достаточно высока. Планируя исследования дневного хода фотосинтеза чая, мы не случайно выбрали метод, обеспечивающий быстрое определение интенсивности фотосинтеза при экспозиции 2 - 3 минуты, так как при этом не допускается перегрева листа в камере. С целью обнаружения кратковременных изменений интенсивности фотосинтеза, определения проводились на одном и том же листе через 10 - 15 минут в течение светового дня. В процессе определения фотосинтеза измеряли: освещенность листьев с помощью люксметра; температуру воздуха; относительную влажность воздуха - аспирационным психрометром Ассмана - МВ-4М и площадь листьев.
Результаты исследования и их обсуждение
В первую очередь нами были определены пороговые величины освещенности и температуры воздуха для процесса фотосинтеза растений чая [1]. На рис. 1 видно, что в пасмурный день, когда максимальная освещенность не превышала 18 тыс. люкс, а температура находилась в пределах 180С - 190С, фотосинтез во второй половине дня (с 14 часов) не превышал 2,0 - 4,0 мгСО2/дм2час. Низкие значения интенсивности фотосинтеза в этот период объясняются действием неблагоприятного водного режима, создавшегося в результате продолжительной засухи (около 6 недель).
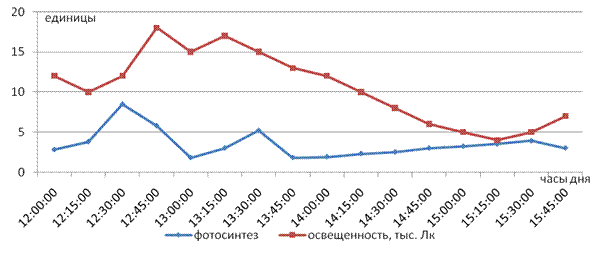
Рис. 1. Влияние низкой освещенности на интенсивность фотосинтеза листьев чая
При этом даже снижение освещенности до 4 тыс. люкс существенно не повлияло на активность этого процесса, чего нельзя сказать о температурном факторе (рис. 2).
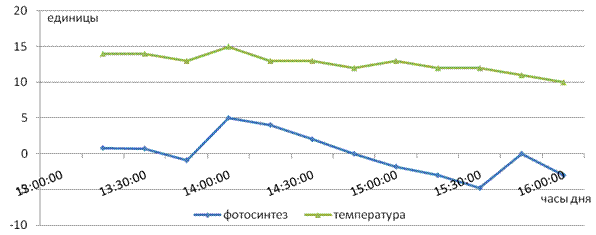
Рис. 2. Влияние пониженной температуры на интенсивность фотосинтеза листьев чая
Так, в дальнейших исследованиях при снижении температуры воздуха (до 10 - 150 С), но высокой освещенности (25 000 - 80 000 Лк) фотосинтез протекал только при температуре выше 140 С (рис. 2). При более низких значениях (10 - 130 С) он сразу же прекращался, и наблюдалось усиление процесса фотодыхания [3]. После установления влияния на процесс фотосинтеза абиотических факторов, мы перешли непосредственно к изучению действия внекорневых обработок микроэлементами. Исследование проводилось на фоне оптимальных значений абиотических факторов. Результаты представлены на рис. 3, отражающем дневной ход фотосинтеза на контрольном варианте. Прежде всего, обращает на себя внимание характер кривой, который указывает, что у чайного растения процесс фотосинтеза протекает очень динамично. В течение светового дня интенсивность фотосинтеза постоянно меняется и имеет «пульсирующий» характер, когда в продолжение даже одного часа при благоприятных погодных условиях усвоения CO2 листьями достигает максимальных значений, а затем резко снижается вплоть до полного прекращения процесса и даже выделения углекислоты в результате активного дыхания.
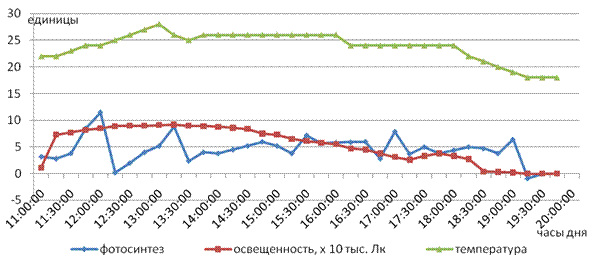
Рис. 3. Дневной ход фотосинтеза листьев чая на контрольном варианте
Особенно активизирует фотосинтез некорневая подкормка растений марганцем. На рис. 4 представлен график дневного хода фотосинтеза листьев чая через неделю после некорневой подкормки, из которого видно, что марганец, быстро проникая в клетки, способствует значительному увеличению интенсивности видимого фотосинтеза, вплоть до максимальных значений (около 20 мгCO2/дм2/час). Одновременно наблюдается усиление дыхательной функции листа, причем, периоды интенсивного поглощения углекислоты довольно равномерно сменяется фазами, когда происходит выделение значительных количеств CO2 в процессе светового дыхания. Причиной таких «пульсаций», по нашему мнению, является накопление в короткое время в хлоропластах большого количества первичных ассимилянтов в период энергичного возрастания интенсивности фотосинтеза и требуется определенное время для «переработки» и оттока метаболитов. В такие моменты интенсивности фотосинтеза резко снижается и начинает превалировать процесс светового дыхания.
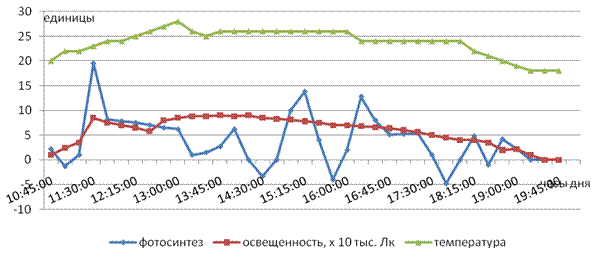
Рис. 4. Влияние марганца на дневной ход фотосинтеза
По мнению некоторых ученых способность хлоропластов быстро увеличивать интенсивность фотосинтеза и также быстро прекращать процесс свидетельствует о высокой физиологической активности фотосинтетического аппарата растений. Что и наблюдается после некорневых подкормок марганцем. Однако активизация хлоропластов со временем снижается и через месяц после обработки растений интенсивность фотосинтеза заметно уменьшается, хотя значительно превосходит этот показатель на контрольном варианте.
Своеобразно выглядит график дневного хода фотосинтеза на варианте с некорневой обработкой растений чая железом (рис. 6).
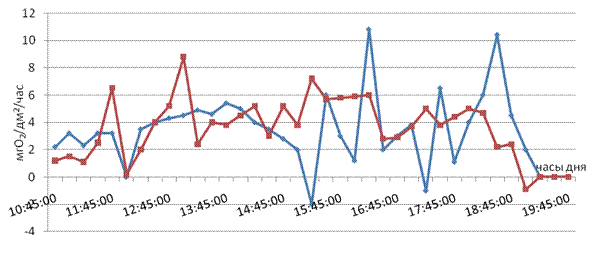
Рис. 6. Влияние железа на дневной ход фотосинтеза
На протяжении исследований мы сталкивались с тем обстоятельством, что на этом варианте растения чая испытывают угнетение жизнедеятельности, что подтверждается данными, например, характеризующими водный режим [1, 4]. Не являются исключением процессы фотосинтеза и дыхания листьев. Из рис. 6 видно, что до 15 часов фотосинтез протекал вяло, интенсивность его составляла 3 - 5 мг СО2 дм/час, хотя освещенность и температура воздуха в это время были благоприятными. И только после 15 часов происходило повышение интенсивности фотосинтеза и фотодыхания листьев, продолжавшееся до конца светового дня, несмотря на снижение потока фотосинтетически активной радиации. В это время наблюдалось увеличение амплитуды и частоты «пульсации» кривой фотосинтеза, что хорошо прослеживается на графике. Однако суммарная продуктивность фотосинтеза на этом варианте, по-видимому, не могла обеспечить функционирования всех органов из-за недостатка энергетического материала, что и вызывает угнетение растений.
Как известно, ответная реакция растений в условиях стресса не однозначна и варьируется в зависимости от вида растения, его устойчивости к стресс-фактору и применяемых агроприемов [5]. Поэтому нами была поставлена задача выявить изменения интенсивности дыхания, происходящие при некорневых подкормках растений чая микроэлементами. На контрастных вариантах опыта с микроэлементами (обработка марганцем и железом) было проведено определение интенсивности дыхания при благоприятном водном режиме и при устойчивом водном дефиците. Результаты определений приведены в таблице 1.
Таблица 1. Интенсивность дыхания листьев чая при внесении микроэлементов
|
Варианты |
благоприятный водный режим |
водный дефицит |
% к исходному |
||
|
мкг О2/г × час |
% |
мкг О2/г × час |
% |
||
|
Контроль |
564,5±12,1 |
10 |
550,1±10,0 |
100 |
- 2,6 |
|
Марганец |
695,2±12,5 |
123,2 |
769,7±9,5 |
139,9 |
10,7 |
|
Железо |
663,1±10,0 |
117,5 |
640,1±5,6 |
116,4 |
- 3,5 |
|
НСР05 |
114,9 |
- |
106,9 |
- |
- |
Представленные данные свидетельствуют о несомненном влиянии некорневых подкормок микроэлементами на интенсивность дыхания. Причем, проявляется закономерность, выявленная нами ранее: обработки марганцем приводят к улучшению физиологического состояния растений чая, что в данном случае проявляется не только в усилении интенсивности синтеза веществ (в процессе фотосинтеза), но и в их окислении, существенно активизируя, таким образом, полный круг метаболических реакций. Усиление интенсивности дыхания на варианте с обработкой железом не существенно и происходит, по нашему мнению, только в силу активизации ферментативных реакций, связанных с окислением веществ. Внесение сернокислого железа приводило к усилению активности каталазы, содержащей этот элемент в качестве кофермента [2]. При водном дефиците интенсивность процесса на контроле и при обработке железом значительно снижается, в то время как обработка марганцем, повышая засухоустойчивость растений, стабилизирует этот процесс - на этом варианте наблюдается некоторое (на 11 %) усиление интенсивности дыхания.
Выводы
Таким образом, можно говорить о том, что некорневые подкормки оказывают значительное влияние на метаболизм клетки, в частности, на процессы ассимиляции и диссимиляции веществ, что, учитывая их роль в повышении стресс-устойчивости, может служить надежным агротехническим приемом, повышающим продуктивность и адаптивные реакции растений чая.
Рецензенты:
- Беседина Т.Д., д.с.-х.н., зав. отделом мелиорации ГНУ ВНИИ цветоводства и субтропических культур Россельхозакадемии, г. Сочи.
- Абильфазова Ю.С., к.б.н., с.н.с. лаборатории физиологии ГНУ ВНИИ цветоводства и субтропических культур Россельхозакадемии, г. Сочи.
- Малюкова Л.С., к.б.н., вед. сотрудник лаборатории агрохимии ГНУ ВНИИ цветоводства и субтропических культур Россельхозакадемии, г. Сочи.
Работа получена 03.11.2011.
Библиографическая ссылка
Белоус О.Г. ВЛИЯНИЕ МИКРОЭЛЕМЕНТОВ НА ИНТЕНСИВНОСТЬ ФОТОСИНТЕЗА РАСТЕНИЙ ЧАЯ В ЗОНЕ ВЛАЖНЫХ СУБТРОПИКОВ РОССИИ // Современные проблемы науки и образования. 2011. № 5. ;URL: https://science-education.ru/ru/article/view?id=4919 (дата обращения: 02.07.2026).