



В данном научном обзоре авторы не ставили перед собой цель специально описывать особенности метаболизма раковых клеток и тем более анализировать особенности метаболизма злокачественных новообразований в сравнении между собой различных типов опухолевых клеток или основных стадий развития опухолевого процесса, включая и степень дифференцированности опухолевых клеток в различных тканях организма, поскольку анализ подобного рода материала предполагает написание отдельной обзорной работы.
Следует отметить, что изучению сигнальных систем, т.е. регуляторных механизмов, влияющих на метаболизм злокачественных новообразований при развитии опухолевого процесса, а также влиянию генетических полиморфизмов на процессы онкогенеза посвящена отдельная работа авторов [1]. Научный обзор, представленный в материале текущей статьи, предлагает вниманию читателей простое наглядное изображение концепции «регуляторно-метаболического старения», которое, по мысли авторов, содержит в себе объяснение многих феноменов, возникающих в процессе старения организмов и одновременно вбирает в себя ключевые теории старения и объясняет такие процессы, как повышение концентрации глюкозы в крови с возрастом (теория старения В.М. Дильмана) или разрушение клеток (теория старения В.А. Эммануэля) свободными радикалами (рисунок).
Интенсивный рост многоклеточных организмов в молодом возрасте или беременность можно рассматривать как своеобразное биохимическое ускорение, которое сопровождается подчинением всех биохимических процессов двум основным целям: биологическому росту и созреванию, а также следующему за ними процессу воспроизводства. Это «ускорение» требует поступления питательных веществ и кислорода растущему организму или плоду в большом количестве и утилизации поступивших в организм соединений по определенным метаболическим путям. Поэтому данный тип метаболизма может быть назван аэробным метаболизмом, или метаболизмом, который чувствителен к молекулярному кислороду.
Процессы гидролитического расщепления углеводов, жиров и белков завершаются образованием пировиноградной кислоты (пирувата) и ацетил-КоА, который окисляется в цикле трикарбоновых кислот с образованием АТФ, СО2 и Н2О [2].
Макроэргические молекулы АТФ расходуются на механическую работу мышечной ткани, на осмотическую работу органов, на создание электрохимического потенциала и проведение по нервным волокнам возбуждения, на процессы поступления глюкозы и аминокислот в клетки (по механизму вторично-активного транспорта) и т.д. [3–5].
Таким образом, при росте организма мы наблюдаем работу идеальной «молекулярной машины».
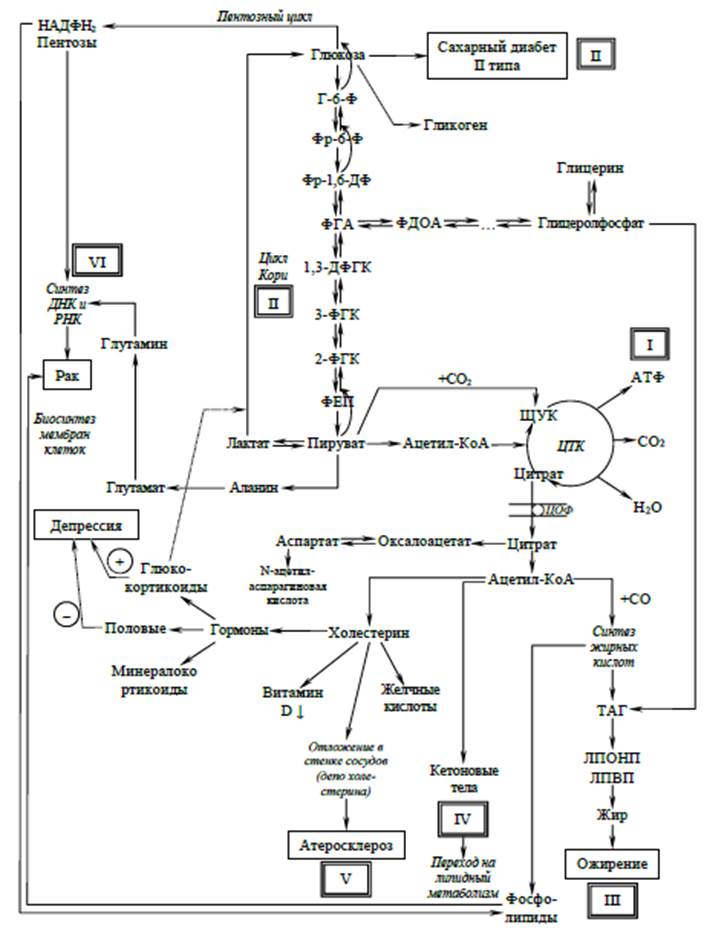
Схема возможных метаболических путей развития патологических состояний
После завершения роста организма наступает, по всей видимости, «анаэробный метаболизм» развития многоклеточных организмов, который является более древним и который становится менее чувствительным через свои сигнальные системы к парциальному давлению кислорода в тканях [6–8], несмотря на достаточное количество кислорода, поступающего в кровь многоклеточных организмов (при этом в условиях «регуляторно-метаболической гипоксии» кислород будет шунтироваться через свободнорадикальные реакции), но становится, по всей видимости, более чувствительным к концентрации углекислого газа в цитозоле клеток, матриксе митохондрий и крови, или, говоря другими словами, включается «обратная метаболическая тяга», к обсуждению которой мы и переходим [9, 10].
Цель исследования: обсуждение участия механизмов аэробного гликолиза и пентозного цикла в неопластических процессах у многоклеточных организмов.
Метаболический дизайн сахарного диабета 2-го типа, или обращенный гликолиз
Обращенный гликолиз хорошо известен биохимикам по реакциям глюконеогенеза, которые сопровождаются сначала превращением лактата в пируват при участии фермента лактатдегидрогеназы (ЛДГ) с последующим карбоксилированием пирувата в оксалоацетат (щавелевоуксусную кислоту, ЩУК) в матриксе митохондрий, превращением ЩУК в малат, выходом яблочной кислоты из митохондрий в цитозоль через специальный белок-переносчик, встроенный в мембрану митохондрий, превращение малата в ЩУК уже при участии цитозольной малатдегидрогеназы, декарбоксилированием ЩУК в фосфоенолпируват (ФЕП) и вхождением ФЕП в «восходящие» гликолитические реакции, которые заканчиваются образованием глюкозы и ее поступлением в кровь как излишнего для клеток метаболита, от которого клетки стремятся избавиться, или, говоря другими словами, глюкоза выступает как своеобразное депо не сгоревшего в ЦТК пирувата. Тем самым теория Дильмана получает твердое биохимическое обоснование. Последующее гликирование гемоглобина эритроцитов, возникновение гипоксического синдрома и развитие сахарного диабета 2-го типа со всеми вытекающими отсюда последствиями являются следствием обращенного гликолиза [11].
Обращенный пентозный цикл
Образуемый в пентозном цикле рибозо-5-фосфат может непрерывно расходоваться на образование пуриновых и пиримидиновых оснований без истощения своего пула, так как пул рибозо-5-фосфата может поддерживаться реакциями обращенного пентозного цикла, а именно реакциями неокислительной стадии пентозного цикла при непосредственном участии метаболитов обращенного гликолиза [12, 13].
В этих реакциях принимают участие такие метаболиты, как 3-ФГА и фруктозо-6-фосфат, образованные в обращенном гликолизе, и собственный метаболит неокислительной стадии пентозного цикла – эритрозо-4-фосфат. При взаимодействии этих метаболитов, 3-ФГА и эритрозо-4-фосфата, образуется седогептулозо-7-фосфат, который может обратимо распадаться на рибозо-5-фосфат и ксилулозо-5-фосфат, а при взаимодействии 3-ФГА и фруктозо-6-фосфата происходит образование эритрозо-4-фосфата и ксилулозо-5-фосфата. В свою очередь, из ксилулозо-5-фосфата возможно образование рибулозо-5-фосфата с последующей его трансформацией в рибозо-5-фосфат.
Таким образом, у обращенного гликолиза появляется еще один «союзник» в виде пентозного цикла, через который возможно «сбрасывать» ненужные метаболиты, такие как 3-ФГА и фруктозо-6-фосфат. Понятно, что побочный продукт в виде рибозо-5-фосфата клетку «мало интересует». Ясно, что подобный «метаболический шунт» соединений, образуемых в гликолизе, создает метаболическую основу для неопластического перерождения клеток [14].
Метаболический дизайн ожирения и атеросклероза, или что происходит с пируватом и ацетил-КоА в цикле трикарбоновых кислот? Гликолиз и пентозофосфатный цикл при аэробном и анаэробном метаболизме, или куда исчезают фосфодиоксиацетон (ФДОА) и фосфоглицериновый альдегид (ФГА), образуемые в гликолизе, и как гликолиз, который был «становым хребтом» для роста организма, становится основой развития ожирения и рака?
Следующий метаболический шунт «ненужного» для клетки пирувата может протекать по классической схеме с образованием ацетил-КоА в матриксе митохондрий с последующим его окислением в ЦТК, но с одной маленькой оговоркой, а именно: цикл трикарбоновых кислот будет «обращенным», т.е. образуемые в нем метаболиты, такие как ЩУК (через аспартат или малат), цитрат (через цитратомыляющий фермент), малат (малат-аспартатный челнок), кетоглутарат (через глутамат), фумарат (через орнитиновый цикл, цикл мочевины), способны покинуть матрикс митохондрии и оказаться в цитозоле клетки. При попадании цитрата в цитозоль из него вновь образуются ЩУК и ацетил-КоА. Последний в цитозоле может участвовать в синтезе жирных кислот и холестерола. Жирные кислоты превратятся в триацилглицериды и отложатся в виде жира, а холестерол может накапливаться в стенках сосудов. Таким образом, «ненужный» пируват превратится в «блудный» пируват, который «обретет покой» в своих физиологических «депо», а именно в жировой ткани (процесс ожирения) и атеросклеротических бляшках (атеросклероз). Кроме того, ацетил-КоА может трансформироваться в кетоновые тела [15], которые будут поступать в кровь, так как клеточные мембраны прозрачны для них, и в условиях дефицита глюкозы из-за обращенного гликолиза (глюкоза выбрасывается из клеток) в кровь организм переходит на липидный обмен при старении, а теория В.М. Дильмана получает еще одно метаболическое подтверждение [16–18].
Превращение аланина в пируват сопровождается образованием глутамата из а-кетоглутарата и дальнейшим образованием глутамина из глутаминовой кислоты. В свою очередь, образование глутамина будет инициировать метаболический путь, участвующий в синтезе пуриновых нуклеотидов, необходимых для синтеза нуклеиновых кислот быстро пролиферирующих клеток [19–21].
Наконец, фосфодиоксиацетон и фосфоглицериновый альдегид из гликолиза могут метаболизироваться в глицеролфосфат с последующим образованием глицерина, принимающего самое непосредственное участие в синтезе фосфолипидов (построение клеточных мембран пролиферирующих клеток) или в синтезе триацилглицеридов (процесс ожирения).
Что происходит с митохондриальными НАДН2 и ФАДН2, образованными в цикле трикарбоновых кислот, или как может трансформироваться работа цитохрома С, который является активным участником процесса апоптоза?
Восстановленные в ЦТК коферменты НАДН2 и ФАДН2 могут при помощи «челноков», малат-аспартатного или глицерофосфатного, переноситься из матрикса митохондрий в цитозоль. При этом фосфодиоксиацетон при участии ФАДН2 восстанавливается в глицеролфосфат, который способен участвовать в синтезе фосфолипидов и триацилглицеролов, а яблочная кислота может при участии «малик-фермента» декарбоксилироваться до пирувата с образованием восстановленной формы НАДФН2, который, в свою очередь, будет востребован в синтезе жирных кислот или в синтезе холестерина.
Восстановленные в ЦТК коферменты ФАДН2 и НАДН2 могут окисляться в дыхательной цепи митохондрий, однако в условиях гипоксии окисление в дыхательной цепи также может иметь обращенный характер.
Смысл обращенного переноса электронов будет состоять в том, что с цитохрома «С» электроны будут направляться не на цитохромоксидазу, а на внешнюю сторону мембраны митохондрий вследствие способности цитохрома «С» мигрировать с внутренней стороны мембраны митохондрий на ее наружную сторону, где электроны будут передаваться в микросомальные цепи с вероятным восстановлением кофермента НАДФН2, который, в свою очередь, может принимать участие в синтезе жирных кислот или в синтезе холестерина [22–24].
Нельзя исключать и предположения о том, что, участвуя в виде «челнока» электронов от дыхательной цепи митохондрий к цепи переноса электронов в микросомах, цитохром «С» «выключается» из участия в процедуре клеточного апоптоза, провоцируя тем самым возникновение мутантных клеточных форм, которые в дальнейшем могут трансформироваться в раковые клетки, неподконтрольные регуляторным воздействиям извне [25–27].
Ацетил-КоА и его участие в развитии сенильного психоза и шизофрении
Известно, что ацетальдегид способен образовывать с серотонином и дофамином соединения в виде сальсонерола, которые могут трансформироваться в морфиноподобные структуры. Вероятность того, что ацетил-КоА, имеющий макроэргическую связь, способен взаимодействовать с серотонином и дофамином, также весьма высока.
Таким образом, намечается еще один путь «депонирования» пирувата в виде морфиноподобных структур, которые, в свою очередь, могут провоцировать увеличение экспрессии опиатных рецепторов на клеточных мембранах и фазовое развитие симптомов шизофрении через последовательное увеличение экспрессии сначала серотониновых, а в последующем – и дофаминовых рецепторов на клеточных мембранах [28–30].
В то же время хорошо известен поставщик метильных групп для ДНК-метилаз. Им является S-аденозилметионин (SАМ), который образуется в цикле гомоцистеина при участии аминокислот серина и глицина. Истощение данного цикла будет приводить к снижению концентрации SАМ и, как следствие, к уменьшению количества метилированных генов, включая и онкогены [31]. Кроме того, снижение концентрации SАМ будет приводить к нарушению синтеза гормона адреналина и медиатора норадреналина, а значит, вызывать нарушение работы адренэргических медиаторных систем организма, что будет способствовать развитию симптомов сенильного психоза [32].
Выводы
По всей видимости, происходит своеобразная метаболическая реверсия гликолиза и пентозного цикла, которая сопровождается возникновением следующих метаболических феноменов: 1) обращенный гликолиз; 2) обращенный пентозный цикл; 3) обращенный цикл трикарбоновых кислот; 4) обращенное митохондриальное дыхание; 5) обращенное микросомальное окисление.
Библиографическая ссылка
Бородулин В.Б., Лосев О.Э., Бобылева Е.В., Никитина В.В., Масальцев А.К., Бородулин Я.В. СТАРЕНИЕ И РАК: ИНВЕРСИЯ ОСНОВНЫХ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ // Современные проблемы науки и образования. 2022. № 1. ;URL: https://science-education.ru/ru/article/view?id=31414 (дата обращения: 04.07.2026).
DOI: https://doi.org/10.17513/spno.31414