



Глобальные проблемы современной цивилизации во многом предопределили возникновение метанаук, принципы которых были спроецированы на образовательную область и обусловили возникновение метапредметов. Поэтому метапредметность присуща не только любой науке, но и любому учебного предмету. Принципы метапредметности особенно актуальны при изучении биологической формы движения материи, которая в скрытом виде содержит в себе физическую и химическую формы ее движения.
Целью настоящего исследования явилась демонстрация эффективности метапредметного подхода, в основу которого положено сопряжение статических и динамических моделей, для истолкования обучающимися сущности фотофизического этапа фотосинтеза в курсе физиологии растений.
Материал и методы исследования. Решению обозначенной проблемы предшествовал анализ публикаций, свидетельствующих об актуальности в современных условиях обучения принципов метапредметного подхода, закрепленных в Федеральных государственных образовательных стандартах, и его эффективности в образовательной практике: М.Н. Ахметовой [1], Т.А. Бахор [2], В.Ф. Бурмакиной [3], Н.В. Громыко [4], С.М. Похлебаева [5], Н.С. Пурышевой [6], Ю.В. Скрипкиной [7], И.А. Третьяковой [8], С.С. Харитоновой [9], А.В. Хуторского [10] и др. В исследованиях данных авторов приведено теоретико-методологическое и методическое обоснование целесообразности и эффективности метапредметного подхода к обучению различных естественно-научных дисциплин (предметов) как в вузе, так и в школе. Наряду с этим многолетний личный опыт авторов позволяет констатировать, что при обучении биологическим дисциплинам в педвузе принципы метапредметного подхода в лучшем случае лишь декларируются, а изучаемый предмет не выстраивается вокруг какой-либо мыследеятельностной организованности.
Результаты исследования и их обсуждение. Государственные образовательные стандарты высшей школы декларируют приоритетность метапредметных основ содержания обучения, которые должны быть конкретизированы и использованы при изучении всех дисциплин данного направления профессиональной подготовки. В качестве такой метапредметной основы выступают, прежде всего, системный и деятельностный подходы. Эти подходы обладают существенным познавательным общенаучным потенциалом, однако, как абстракции высокого уровня, они обедняют существующую реальность, что детерминирует их конкретизацию.
Конкретизация принципов системного и деятельностного подходов очень четко просматривается при построении моделей как на эмпирическом, так и теоретическом уровне. Данные методологии тесно связаны между собой, и поэтому некоторые авторы считают, что моделирование можно одновременно рассматривать как разновидность и конкретизацию системного и деятельностного подходов. Теоретические аспекты данных методологий разработаны достаточно хорошо, однако их применение обучаемыми в школах и вузах оставляет желать лучшего. Особенно это касается моделирования физиолого-биохимических процессов при изучении биологических дисциплин. Данные процессы не подлежат прямому наблюдению, поэтому их сущность наиболее полно можно отразить в наглядных моделях.
Моделирование обладает огромной эвристической силой, что находит отражение в создании у обучаемых наглядного обобщенного образа моделируемого объекта, возникающего при конструировании модели. Кроме того, построение моделей способствует взаимодействию эмпирического и теоретического уровней познания, мышления с чувственностью, не наглядных элементов с наглядными, что соответствует требованиям современной науки и облегчает понимание формальных теорий. Метод моделирования обеспечивает стратегию познания, направляет мышление к достижению цели кратчайшим путем.
По мнению Л.М. Фридмана, школьники, изучающие естественные дисциплины, имеют «весьма смутные и ограниченные представления о моделировании и моделях». Вместе с тем, отмечает данный автор, психологические исследования свидетельствуют, что обучение школьников моделированию как общенаучному методу познания детерминирует формирование у них научного мировоззрения, обогащает их методологический аппарат и существенно изменяет их отношение к учебным предметам, к учению. При этом их учебная деятельность становится более осмысленной и продуктивной [11, с. 89].
Исследования, проведенные М.Ю. Королевым, свидетельствуют, что существующие методики обучения студентов естественно-научных направлений подготовки в педвузах методу моделирования также не являются достаточными для обеспечения должного уровня профессиональной компетентности будущих педагогов [12].
Потребность в моделировании физиолого-биохимических процессов особенно стала актуальной при выявлении их сущности на молекулярном и электронном уровне. Моделирование как метод обучения особенно востребован при интерпретации физических и химических явлений, лежащих в основе таких важнейших физиолого-биохимических процессов, как фотосинтез и дыхание, которые являются основными звеньями углеводного метаболизма растительной клетки.
Исторически важную роль в изучении сущности энергетических преобразований, имеющих место на фотофизическом этапе, сыграл метод моделирования, использование которого при обучении физиологии растений также должно стать приоритетным.
Общая модель энергетических преобразований энергии квантов света на фотофизическом этапе представлена в учебнике по физиологии растений, автором которого является В.В. Полевой (рисунок 1).
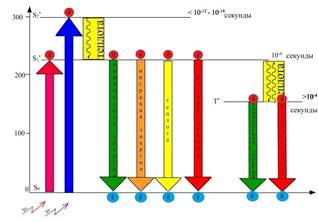
Рис. 1. Энергетическое состояние молекулы хлорофилла и различные пути использования энергии электронного возбуждения
Однако практика свидетельствует, что большинство обучающихся испытывают значительные трудности в интерпретации этой модели и установлении логических связей между ее отдельными элементами. Это послужило основанием для разработки еще 7 (дополнительных) статических моделей, содержание которых фиксирует отдельные энергетические преобразования, которые имеют место на этом первоначальном этапе фотосинтеза.
На рисунке 2 внимание обучающихся акцентировано на то, что кванты красного и сине-фиолетового света поглощаются электронами молекулы хлорофилла, находящимися в основном синглетном состоянии (S0) и обладающими минимальным запасом энергии.
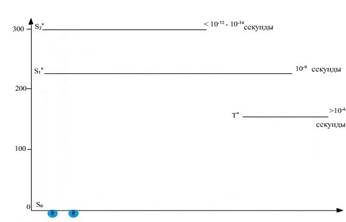
Рис. 2. Положение электронов в молекуле хлорофилла, находящейся в основном синглетном (S0) энергетическом состоянии
На рисунке 3 зафиксировано, что электрон, поглотивший кванты красного света, перешел на более высокий энергетический уровень (S1). Для снятия вопроса студентов, почему возбужденный светом электрон перешел только на первый уровень (S1), рисунок дополнен физической формулой, отражающей обратную зависимость между длиной волны и энергией кванта света.
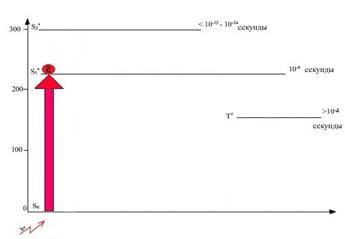
Рис. 3. Положение электрона в молекуле хлорофилла после поглощения ею кванта красного света. (Молекула хлорофилла находится в первом синглетном возбужденном состоянии – S*1)
Данные о длинах волн студенты получают, используя рисунок 4 (шкалы солнечного спектра), производя подсчеты, они убеждаются, что красные лучи являются длинноволновыми и поэтому их кванты обладают меньшей энергией.
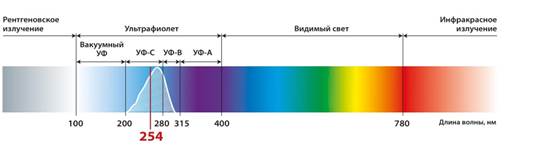
Рис. 4. Шкала электромагнитных излучений
На рисунке 5 зафиксирован момент перехода электрона (находящегося в основном синглетном состоянии) (S0), поглотившего квант синего света, на более высокий энергетический уровень (S2). На этом же рисунке также имеется и формула, позволяющая объяснить данный переход.
На последующих моделях отражены различные моменты (пути) преобразования и использования энергии возбужденных электронов молекулы хлорофилла. Судьба энергии электрона зависит от его времени жизни на разных энергетических уровнях. Существует общая закономерность, согласно которой, чем выше энергетический уровень, на котором находится электрон, тем меньше временной период, в течение которого он может там находиться. Кроме того, для преобразования одной формы энергии в другую или ее использования необходим определенный промежуток времени.
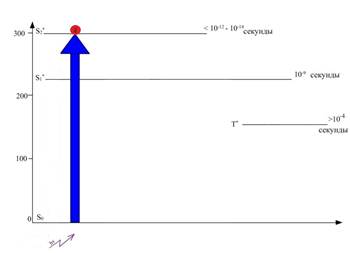
Рис. 5. Положение электрона в молекуле хлорофилла после поглощения ею кванта синего света. (Электрон переходит на более высокую энергетическую орбиталь – S*2)
Так, на рисунке 6 зафиксировано, что время жизни электрона S2 орбитали - самое наименьшее и составляет всего лишь 10 -12 – 10 -14 секунды. Это время настолько мало, что на его протяжении энергия электронного возбуждения не может быть использована. Через короткий промежуток времени электрон переходит на более низкий энергетический уровень (S1), сопровождающийся некоторой потерей энергии, выделяющейся в виде теплоты. По величине энергии данный электрон сравнялся с энергией электрона, поглотившего квант красного света.
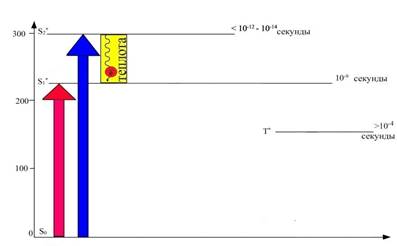
Рис. 6. Безызлучательный переход электрона с более высокой энергетической орбитали – S*2, на более низкую энергетическую орбиталь – S*1, сопровождающийся выделением
энергии в виде теплоты
Дальнейшая судьба электронов, находящихся в первом синглетном состоянии, может быть различной. Они могут переходить с изменением спина электрона на триплетный (Т1) уровень, сопровождающийся некоторой потерей энергии, выделяющейся в виде теплоты. Либо из возбужденного первого синглетного (S1) и триплетного (Т1) состояний может переходить в основное (S0).
При переходе электронов с S1 на S0 - энергетический уровень энергия электрона может быть: передана другим, не возбужденным молекулам хлорофилла (миграция энергии), выделена в виде теплоты, света - флуоресценции (испускание света с измененной длиной волны), преобразована в химическую форму энергии (рисунок 7). При этом важно отметить, что в последнем преобразовании электрон не переходит в основное синглетное состояние, а покидает эту орбиталь и уходит в электронтранспортную цепь, где его энергия используется для синтеза таких энергетических эквивалентов, как АТР и NADPH. Чтобы хлорофилл смог принять следующую порцию энергии, он должен восстановиться, т. е. вернуть себе электрон. Свободная орбиталь (S0) будет заполнена электроном воды, образующимся в процессе ее фотоокисления.
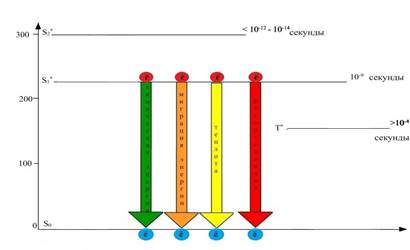
Рис. 7. Пути возвращения молекулы хлорофилла в основное состояние: с выделением теплоты, с испусканием света в виде флуоресценции, с миграцией энергии к невозбужденным молекулам хлорофилла, с использованием энергии на фотохимические реакции
Как уже было отмечено выше, с первого синглетного (S1) состояния электрон может переходить на более низкий энергетический уровень (Т1) - триплетный, сопровождающийся некоторой потерей энергии, выделяющейся в виде теплоты (рисунок 8).
С триплетного уровня (Т1) электрон может переходить в основное синглетное состояние (S0), а его энергия в этом случае может испускаться в виде фосфоресценции (более длительного свечения) или потрачена на химическую работу (рисунок 9). В последнем случае электрон покидает орбиталь триплетного уровня (Т1) и поступает в ЭТЦ, которая встроена в мембраны тилакоида хлоропласта, где его энергия будет использована для синтеза АТР и NADPH. В этом случае S0 − орбиталь также будет заполнена электроном молекулы воды, которая является конечным донором электронов в ЭТЦ.
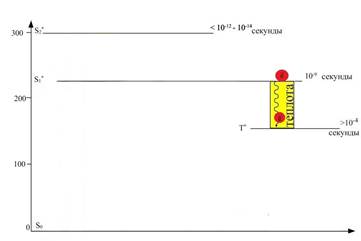
Рис. 8. Потеря энергии в виде теплоты при переходе молекулы хлорофилла из первого
синглетного возбужденного состояния (S*1) в триплетное состояние (Т*)
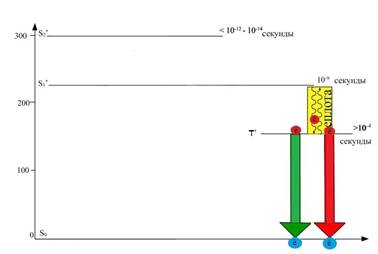
Рис. 9. Потеря энергии при переходе молекулы хлорофилла из триплетного возбужденного состояния (Т*) в основное (S*0) в виде фосфоресценции (слабого свечения), или использовании энергии на фотохимические реакции
По мнению И.Д. Зверева и А.Н. Мягковой, «... в работе с экранными пособиями в качестве источника знаний выступает сочетание слова учителя и изобразительной наглядности, статистических и динамических изображений» [13, с. 136]. Внедрение новых информационных технологий позволяет развить эту идею авторов и применить в образовательном процессе для интерпретации фундаментальных теорий статические и динамические flash–модели, в комплексе. Такое моделирование, по-видимому, можно назвать сопряженным. Оно позволяет имитировать физиолого-биохимические процессы на молекулярном и даже электронном уровне, проводить виртуальные наблюдения за этими процессами, останавливать их для детального рассмотрения отдельных этапов и пр. При этом перед обучающимися открываются большие познавательные возможности, запрограммированно инициируются чувственные и интеллектуальные эмоции, которые предопределяют внутреннюю мотивацию к изучению биологических объектов и явлений.
Заключение. Таким образом, сопряжение статических и динамических моделей при изучении физиолого-биохимических процессов является весьма эффективным методом познания, так как позволяет достаточно быстро перейти от наглядно-образного типа мышления к обобщенно-образному а от него к понятийному типу мышления. Сопряженное моделирование как метод познания может использоваться как в иллюстративном, так и в поисковом плане - это зависит от дидактических целей, методологического потенциала разработанных моделей. Использование блока дополнительных статических моделей, отражающих последовательность отдельных (конкретных) энергетических преобразований, в сочетании с динамической моделью, демонстрирующей фотофизический процесс в динамике, позволяет обучающимся усвоить материал курса физиологии растений, убедиться в эффективности использованного данного метапредметного подхода обучения и в последующем применить его в их профессиональной деятельности.
Библиографическая ссылка
Похлебаев С.М., Манжукова Л.Ф., Третьякова И.А. СОПРЯЖЕНИЕ СТАТИЧЕСКИХ И ДИНАМИЧЕСКИХ МОДЕЛЕЙ КАК МЕТАПРЕДМЕТНАЯ ОСНОВА ДЛЯ ПОНИМАНИЯ СУЩНОСТИ ФОТОФИЗИЧЕСКОГО ЭТАПА ФОТОСИНТЕЗА // Современные проблемы науки и образования. 2020. № 6. ;URL: https://science-education.ru/ru/article/view?id=30369 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/spno.30369