



В настоящее время колоректальный рак (КРР) – одно из наиболее распространенных злокачественных новообразований в мире. Ежегодно регистрируется около 1 000 000 новых случаев этого заболевания и более 700 000 летальных исходов. За последние 10 лет в России заболеваемость КРР значительно увеличилась, и по числу новых случаев и числу умерших больных он уступает лишь раку легкого, желудка и молочной железы [1].
Хирургические методы и лучевая терапия в интеграции с химиотерапией являются основными направлениями лечения КРР. Один из вариантов предоперационного лечения – это короткий курс лучевой терапии, который проводится на первичную опухоль и зону регионарного метастазирования за 5 фракций с разовой очаговой дозой (РОД) 5 Гр до СОД (суммарной очаговой дозы) 25 Гр, что изоэффективно 40 Гр. Большое влияние на эффективность подобной терапии оказывает исходная радиорезистентность опухолевых клеток, зависящая от их молекулярно-генетических особенностей [1]. К подобным особенностям можно отнести показатель копийности генов (Copy Number Variation (CNV)) – вид генетического полиморфизма, результатом которого может явиться снижение или повышение числа копий определенного гена и, следовательно, пониженная или повышенная экспрессия продукта гена-белка или некодирующей РНК [2, 3].
Целью данной работы явилось исследование влияния показателя копийности генетических локусов, ответственных за регуляцию репарации ДНК, клеточного цикла и апоптоза (AKT, ATM, BRIP, BRCA1, BRCA2, CDK1, CDKN1B, CCND1, CCND3, EXO1, FGFR2, HIST1, H2AX, KU70, PTEN, RAD50, RAP80, RIF1, RNF168, TOPB1, TP53, XRCC4, BAX, CASP8, CASP3, CASP9, MDM2, BCL2, RBBP8, EP300, LIG4, C-FLIP), на радиорезистентность клеток линии HT-29 в условиях воздействия стандартных доз лучевой терапии 5 и 7 Гр на протяжении 5 дней.
Материалы и методы исследования. Для модельного эксперимента использовалась культура клеток человека HT-29 (аденокарцинома толстой кишки). Манипуляции с клеточной линией проводились в ламинарном шкафу II степени биологической защиты. Культивирование осуществлялось в инкубаторе CB 150 (Binder, Германия) в стерильных плоскодонных флаконах, содержащих среду RPMI-1640 c 10%-ной фетальной телячьей сывороткой и концентрацией гентамицина 50 мкг/мл в условиях 5% CO2 и 95% влажности при 370C. Облучение проводилось на линейном ускорителе Novalis TX (Varian, США) (5 раз через каждые 24 часа). Использовались варианты фракционирования РОД=5 (СОД=25 Гр, изоэффективная СОД=37,5 Гр) и РОД=7 Гр (СОД=35 Гр, изоэффективная СОД=49,6 Гр) [1].
Оценку общего количества клеток, а также соотношения живых и мертвых клеток проводили в камере Горяева (0,4%-ный раствор трипанового синего). На пятый день облучения, после микроскопирования (DM IL LED Fluo (Leica, Германия)), клеточную линию снимали с подложки флаконов (3 контрольных флакона и 6 флаконов для дозы 5 Гр и 7 Гр соответственно) путем трипсинизации раствором Трипсин/Версена [1]. После этого клеточную массу отмывали от среды фосфатно-солевым буферным раствором Дюльбекко (DPBS) («Биолот», Россия) и осаждали центрифугированием.
Из клеточных препаратов с использованием наборов ДНК-Сорб-В («АмплиСенс», Россия) проводилась экстракция ДНК.
Определение относительной копийности 32 генетических локусов (AKT, ATM, BRIP, BRCA1, BRCA2, CDK1, CDKN1B, CCND1, CCND3, EXO1, FGFR2, HIST1, H2AX, KU70, PTEN, RAD50, RAP80, RIF1, RNF168, TOPB1, TP53, XRCC4, BAX, CASP8, CASP3, CASP9, MDM2, BCL2, RBBP8, EP300, LIG4, C-FLIP) проводили методом Real-Time qPCR (RT-qPCR). Принцип метода заключается в одновременной амплификации гена-мишени и референсного гена в опытной и контрольной пробах. Вывод об изменении дозы гена делается на основании анализа соотношения сигналов, продуцируемых ампликонами изучаемой и референсной последовательностей [2].
Количественная RT-PCR амплификация проводилась с использованием термоциклера Bio-Rad CFX96 (Bio-Rad, USA) в соответствии с инструкциями производителя по следующей программе: 95°C 5 мин и 40 циклов при 95°C 10 сек, 58°C 30 сек (чтение оптического сигнала FAM) и 72°C 30 сек. Каждые 20 мкл ПЦР-смеси для анализа содержали 2,4 нг геномной ДНК, 0,2 mM dNTP’s, по 600 нМ прямого и обратного праймеров для референсного гена (GAPDH) или гена-мишени, 2,5 mM MgCl2, 1X ПЦР-буфер, 0,1 u/µl SynTaq ДНК-полимеразы с ингибирующими активность фермента антителами («Синтол», Россия) [2]. В качестве красителя использовали SYBR Green I. Амплификация каждой из проб осуществлялась в трех повторностях. Первичные данные RT-qPCR получали с использованием программного обеспечения Bio-Rad CFX Manager ver 3.1.
Прямые и обратные праймеры были разработаны нами с использованием базы данных NCBI GenBank. Генетические локусы GAPDH, ACTB и B2M применяли в качестве референсных для нормализации полученных показателей количественной RT-qPCR.
Статистический анализ выполняли в прикладном пакете программ STATISTICA 10.0. Нормальность распределения показателей оценивали с помощью критерия Шапиро–Уилка (n<50). Для оценки различий использовали критерий Манна–Уитни для порогового уровня статистической значимости р<0,05. Для построения Heatmap и проведения кластерного анализа (Hierarchical Clustering, Euclidean distance) применяли собственные скрипты в среде R (R-Studio 8.10.173.987).
Результаты исследования и их обсуждение
После 5-дневного облучения на линейном ускорителе Novalis TX культуры клеток HT-29 в дозах 5 и 7 Гр обнаружено статистически значимое снижение общего количества клеток на 56 и 72% соответственно (р<0,05) (рис. 1). При этом СОД=35 Гр (5 х РОД=7 Гр) оказала более выраженный эффект на элиминацию опухолевых клеток, чем СОД=25 Гр (5 х РОД=5 Гр).
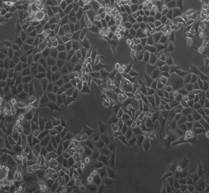

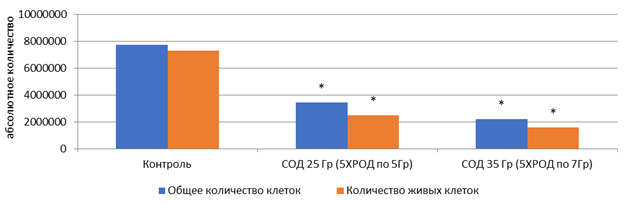
Рис. 1. Микрофотография клеточного монослоя HT-29 после 5-го дня облучения (слева направо: контроль – СОД 25 Гр (изоэффективная доза 37,5 Гр) – СОД 35 Гр (изоэффективная доза 49,6 Гр)), общее количество и количество живых клеток.
* – статистически значимые отличия (р<0,05) относительно контроля
После облучения только определенный пул из оставшегося количества клеток НТ-29 сохранил жизнеспособность. Доля таких клеток для СОД 25 Гр составила 32,3%, а для СОД 35 Гр – 20,4% от изначального количества клеток, взятых в эксперимент. Наблюдаемый эффект связан с повышенной радиорезистентностью этих клеток относительно других клеток линии HT-29, вероятно, благодаря определенным молекулярно-генетическим особенностям. Известно, что ионизирующее излучение вызывает повреждения ДНК в опухолевых клетках, индуцируя двухцепочечные разрывы [4]. Но некоторые опухолевые клетки способны использовать активировать системы репарации двухцепочечных разрывов для преодоления цитотоксичности от излучения, а также иные сигнальные пути, не являющиеся компонентами репарационных систем клетки [5].
В результате 5-дневного эксперимента по облучению клеток HT-29 нами были получены данные о дифференциальном показателе копийности 32 генов в интактных (контрольных) и облученных клетках (рис. 2).
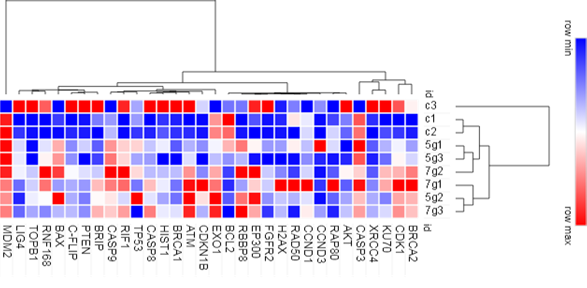
Рис. 2. Heatmap и кластерный анализ дифференциальной копийности 32 генов в интактных (контрольных) и облученных клетках НТ-29
По особенностям уровня копийности в интактных и облученных образцах в результате анализа было выделено 4 основных кластера генов: 1 – (BRCA2, CDK1, KU70, XRCC4, CASP3), 2 – (AKT, RAP80, CCND3, CCND1, RAD50, H2AX, FGFR2, EP300, RBBP8, BCL2), 3 - (EXO1, CDKN1B, ATM, BRCA1, HIST1, CASP8, TP53, RIF1, CASP9), 4 – (BRIP, PTEN, C-FLIP, BAX, RNF168, TOPB1, LIG4, MDM2). При этом сами интактные и облученные образцы по схожести копийности генов объединяются в кластеры 1 (с1,с2), 2 (5g1, 5g3, 7g2) и 3 (7g1, 5g2, 7g3), которые далее образуют кластеры 4 (кластер 1 и 2), 5 (кластер 4 и 3) и 6 (кластер 5 и c3). Сходство по уровню копийности некоторых генов в интактных и облученных образцах на разных уровнях кластеризации можно объяснить изначальным наличием в линии HT-29 субпопуляции клеток с определенным уровнем копийности рассматриваемых генов. Пониженный или повышенный уровень копийности этих генов в условиях лучевой терапии, очевидно, может обеспечить выживание субпопуляции этих клеток.
После нормализации показателей копийности в облученных клетках относительно интактных были получены данные, отраженные на рисунке 3. В клетках НТ-29, подвергнутых облучению СОД 35 Гр, статистически значимо была повышена копийность генов BRCA2, H2AX, CASP9 и RBBP8: в 2,5 раза; 3,2 раза; 1,6 раза и 1,6 раза соответственно (р<0,05), и снижена копийность гена BCL2 в 3,6 раза (р<0,05) относительно интактных клеток. В клетках НТ-29, подвергнутых облучению СОД 25 Гр, статистически значимо была повышена копийность только 2 генов – CASP9 и RBBP8 в 1,5 и 1,4 раза соответственно (р<0,05).
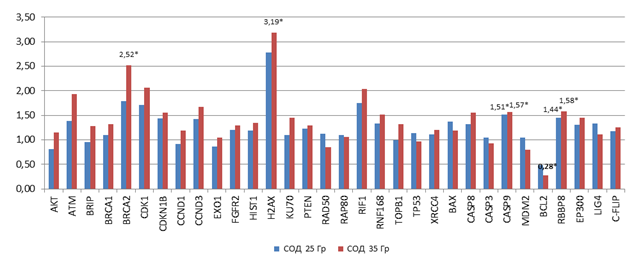
Рис. 3. Относительная копийность генов в клетках НТ-29 с повышенной радиорезистентностью (после облучения в дозах 5 и 7 Гр в течение 5 дней).
*– статистически значимые отличия относительно интактных (контрольных) клеток (р<0,05)
Анализ сигнальных путей, в которых задействованы гены BCL2, BRCA2, H2AX, CASP9 и RBBP8, с помощью онлайн-сервиса GIANT (Genome-scale Integrated Analysis of gene Networks in Tissues, http://giant.princeton.edu/) [6] позволил визуально представить взаимодействия данных генов, а также силу этих взаимодействий (рис. 4).
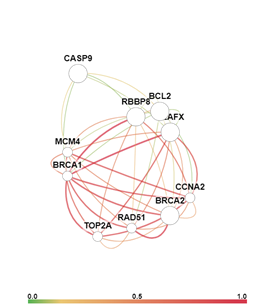
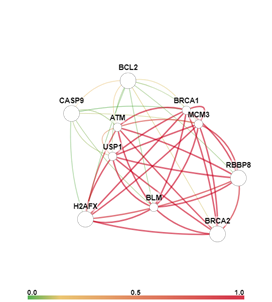
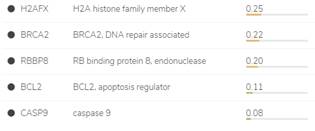

Рис. 4. Сеть взаимодействия и индекс взаимосвязи генов BCL2, BRCA2, H2AX, CASP9 и RBBP8 в ткани кишки (слева) и в ткани кишки в условия воздействия ионизирующего излучения (справа)
BCL2, BRCA2, H2AX, CASP9 и RBBP8 являются компонентами различных сигнальных каскадов нормальных и опухолевых клеток, и изменение их экспрессии, опосредованное изменением копийности, приводит к изменению экспрессии целого ряда других генов [7] (рис. 5).
С использованием «Алгоритма сетевой интеграции с несколькими ассоциациями» (Gene Multiple Association Network Integration Algorithm) были установлены взаимодействия между генами BCL2, BRCA2, H2AX, CASP9, RBBP8 и APAF1 (apoptotic peptidase activating factor 1), RAD51, APPL1, SPO11 (initiator of meiotic double stranded breaks), FKBP8, RAD50 (double strand break repair protein), MRE11A (double strand break repair nuclease), BID (BH3 interacting domain death agonist), BIRC5 baculoviral IAP repeat containing), TP53BP1, BIK (BCL2 interacting killer), BAX (BCL2 associated X protein), SHFM1 (split hand/foot malformation (ectrodactyly) type 1), ATM (serine/threonine kinase) и ENY2 (рис. 5). «Алгоритм сетевой интеграции с несколькими ассоциациями» предсказывает функцию гена в составе сложной сети из множества генов, используя разновидность алгоритма «распространения меток гауссовых полей» (один из алгоритмов машинного обучения). Алгоритм присваивает оценку (значение дискриминанта) каждому узлу-точке построенной сети. Эта оценка отражает вычисленную силу связи. Так, физические взаимодействия между продуктами данных генов составили 67,6%, со-экспрессия – 13,5%, со-локализация – 6,2%, взаимодействия в рамках общих сигнальных путей – 4,4%, генетические взаимодействия – 1,4% и общие белковые домены – 0,5% от всех вычисленных взаимодействий [8].
Упаковка эукариотической ДНК в виде хроматина представляет собой преграду для всех процессов, которые требуют привлечения ферментов к определенным сайтам ДНК. Для репарации ДНК после облучения хроматин должен быть ремоделирован (деконденсирован) [9].
Ген H2AX кодирует гистоновый белок из семейства H2A. В ответ на двухцепочечные разрывы в ДНК, вызванные ионизирующим излучением, H2AX становится фосфорилированным по серину в положении 139 (γH2AX). За его фосфорилирование ответственны киназы семейства PI3 (Ataxia telangiectasia mutated, ATR и DNA-PKcs), особенно ATM (рис. 4). Из-за этой модификации (фосфорилированной формы гистона) ДНК становится менее конденсированной, освобождается место для присоединения белковых комплексов, необходимых во время репарации.
Когда в ДНК происходит двухцепочечный разрыв, возникает последовательность событий, при которых изменяется H2AX. В течение одной секунды после двухцепочечного разрыва белок HP1-бета фосфорилируется и высвобождается из хроматина [10].
Высвобождение HP1-бета приводит к динамическому изменению в структуре хроматина. Это изменение в структуре хроматина в свою очередь способствует фосфорилированию H2AX с помощью ATM, ATR и DNA-PK [10], что приводит к образованию γH2AX (через 20 секунд после облучения клеток). Затем MDC1 связывается с γH2AX. Убиквитинлигазы RNF8 и RNF168 связываются с комплексом γH2AX/MDC1, убиквитилируя другие компоненты хроматина. Это позволяет BRCA1 присоединиться к хроматину, модифицированному γH2AX/MDC1 [11] Среди других белков, которые собираются на участке γH2AX-модифицированного хроматина, нужно выделить комплекс MRN (белковый комплекс, состоящий из Mre11, Rad50 и Nbs1), RAD51 и киназу АТМ [12].
Продукт гена BRCA2 также необходим для восстановления поврежденной ДНК. BRCA2 связывает одноцепочечную ДНК и напрямую взаимодействует с рекомбиназой RAD51, чтобы стимулировать важный этап гомологичной рекомбинации. Перемещение RAD51 к двухцепочечному разрыву ДНК требует образования комплекса BRCA1-PALB2-BRCA2 [13].

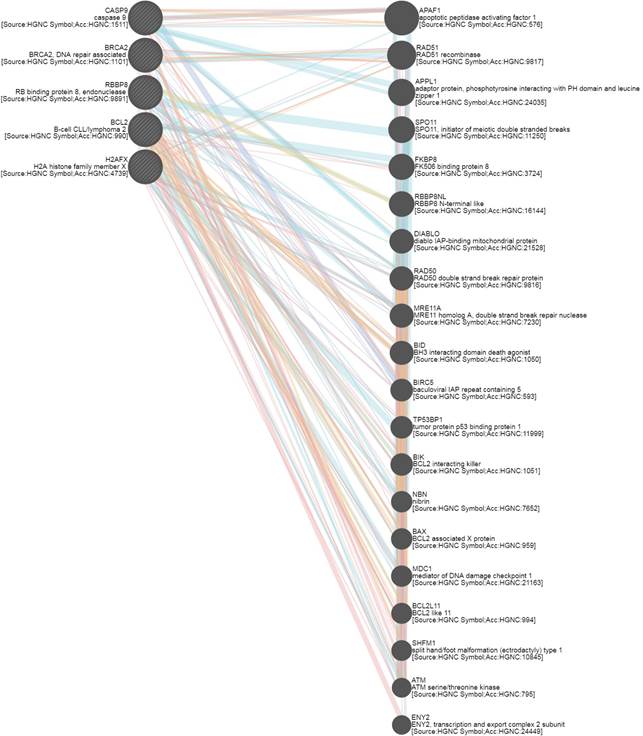
Рис. 5. Взаимодействия генов, вычисленные с помощью GeneMANIA
(Gene Multiple Association Network Integration Algorithm, Алгоритм сетевой интеграции с несколькими ассоциациями) [8]
RBBP8 кодирует белок, который регулирует пролиферацию клеток. Этот белок образует комплексы с транскрипционным ко-репрессором CTBP, а также модулирует функции BRCA1 в регуляции транскрипции и репарации ДНК [14].
Соответственно повышенная копийность генов BRCA2, H2AX и RBBP8 в отдельных клонах клеточной линии НТ-29 дает им преимущество в виде более эффективно действующей системы репарации ДНК и обеспечивает выживание при воздействии лучевой терапии.
Продукты генов BCL2 и CASP9 вовлечены в регуляцию апоптоза. BCL2 подавляет апоптоз во многих клеточных системах, контролируя проницаемость митохондриальной мембраны, ингибирует каспазы за счет предотвращения выхода цитохрома C из митохондрий и за счет связывания фактора, активирующего апоптоз, – APAF1. CASP9 кодирует инициаторную каспазу, критическую для запуска апоптоза. Апоптотические сигналы вызывают высвобождение цитохрома С из митохондрий и активацию APAF-1, который затем расщепляет про-фермент каспазы-9, переводя его в активную форму [15].
Можно предложить, что выжившие клоны клеточной линии НТ-29 изначально могли обладать более мощной системой репарации ДНК (повышенной копийностью и, вероятно, экспрессией генов BRCA2, H2AX и RBBP8), а также более эффективной системой регуляции апоптоза: повышенной копийностью проапоптозного гена CASP9 и сниженной копийностью антиапоптозного гена BCL2.
Заключение. Проведенное модельное исследование позволило установить, что линия клеток НТ-29 исходно гетерогенна по копийности некоторых генов, и пятидневная лучевая терапия при РОД 5 и 7 Гр с использованием ускорителя Novalis TX приводит к селективному выживанию 32,3% и 20,4% клеток соответственно, обладающих повышенной копийностью генов BRCA2, H2AX, RBBP8, CASP9 и сниженной копийностью гена BCL2.
Библиографическая ссылка
Кутилин Д.С., Кошелева Н.Г., Гусарева М.А., Потемкин Д.С., Полуэктов С.И., Дашков А.В., Каймакчи Д.О., Носов В.А., Газиев У.М., Легостаев В.М. КОПИЙНОСТЬ ГЕНОВ КАК ФАКТОР РАДИОРЕЗИСТЕНТНОСТИ КЛЕТОК АДЕНОКАРЦИНОМЫ ТОЛСТОЙ КИШКИ ЛИНИИ HT-29 // Современные проблемы науки и образования. 2019. № 5. ;URL: https://science-education.ru/ru/article/view?id=29224 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/spno.29224