



Бедренная кость человека как часть системы опорно-двигательного аппарата организма с бипедальной локомоцией формируется под действием сил, обеспечивающих вертикальное положение тела при передвижении в пространстве. Это взаимосвязанная система рычагов, расположенных в двух крупных суставах – тазобедренном и коленном. Форма бедренной кости, степень развития ее апофизов, изогнутость диафиза и углы, образованные между осями кости, отражают условия функционирования биомеханической системы нижних конечностей и формируются в процессе жизни человека [1]. В связи с постоянным увеличением числа дегенеративно-дистрофических заболеваний крупных суставов нижних конечностей, возрастающей потребностью в реэндопротезировании тазобедренных суставов [2] строение бедренной кости должно быть достаточно серьезно изучено, однако до сих пор отсутствует четкое представление о взаимосвязях между линейными и угловыми размерами ее анатомических образований. Выявить структуры кости человека, наиболее подверженные изменениям при патологии, невозможно без глубокого сравнительно-анатомического анализа структурной организации бедренных костей животных с различными типами опоры на конечность и человека, что и определило высокую актуальность нашего исследования.
Цель работы: изучить структурную организацию бедренной кости человека и животных с различными типами опоры на заднюю конечность.
Материалы и методы исследования
Настоящее исследование проведено с соблюдением всех этических и правовых норм, о чем свидетельствует разрешение регионального этического комитета Курского государственного медицинского университета (протокол заседания РЭК № 5 от 30 июня 2018 г.).
Для исследования были отобраны бедренные кости фалангоходящих животных – Bos taurus taurus, L.1758 (40 правых и 40 левых); пальцеходящих – Canis lupus familiaris, L.1758 (по 39 правых и левых); стопоходящих – Oryctolagus cuniculus, L.1758 (по 40 правых и левых) и современного человека (77 правых и 77 левых). Кости отличались полным синостозированием эпифизов и не имели признаков патологии. Определение половой принадлежности не проводилось.
Дизайн исследования включал остеометрию гомологичных структур на бедренных костях [3], статистическую оценку выборок классическими методами [4], создание математических моделей структурной организации бедренных костей человека и животных с учетом принадлежности к стороне тела.
Методика остеометрии гомологичных структур бедренных костей представлена в таблицах 1–3.
Таблица 1
Остеометрия структур проксимального эпифиза
|
Наименование структуры |
Расшифровка измерений |
|
ШПЭ |
Наибольшее расстояние между головкой и большим вертелом |
|
ØГС |
Сагиттальный диаметр головки или переднезадний размер головки, измеренный в горизонтальной плоскости |
|
ØГВ |
Вертикальный диаметр головки или верхненижний размер головки, измеренный во фронтальной плоскости |
|
ØШС |
Сагиттальный диаметр шейки или переднезадний размер шейки, измеренный в горизонтальной плоскости |
|
ØШВ |
Вертикальный диаметр шейки. Расстояние, измеренное между максимально удаленными верхней и нижней точками шейки кости |
|
ПДШ |
Передняя длина шейки бедренной кости – расстояние между краем головки спереди до середины межвертельной линии |
|
ЗДШ |
Задняя длина шейки – расстояние между краем головки спереди до середины межвертельного гребня |
|
ВДШ |
Верхняя длина шейки бедренной кости – расстояние между большим вертелом и краем головки сверху |
|
НДШ |
Нижняя длина шейки – расстояние между краем головки до малого вертела |
|
МБР |
Расстояние между вершинами большого и малого вертела бедренной кости |
|
ДШУ |
Диафизарно-шеечный угол |
|
АНТ |
АНТ – угол анте- (ретро)версии шейки кости или угол отклонения оси шейки кпереди или кзади от фронтальной плоскости, проведенной по задней поверхности мыщелков |
|
ТОР |
ТОР – угол сочленения шейки бедренной кости с диафизом в сагиттальной плоскости |
Таблица 2
Остеометрия структур диафиза
|
Наименование структуры |
Расшифровка измерений |
|
Дг |
Наибольшая длина кости, измеренная между максимально удаленными точками на нижней поверхности медиального мыщелка и наивысшей точкой на головке |
|
Дбв |
Наибольшая длина кости, измеренная между максимально удаленными точками на нижней поверхности медиального мыщелка и наивысшей точкой на большом вертеле |
|
ØДп |
Поперечный диаметр диафиза – расстояние между боковыми точками на середине длины кости во фронтальной плоскости |
|
ØДс |
Сагиттальный диаметр диафиза – переднезаднее расстояние, измеренное на середине длины кости |
|
УНД |
Угол наклона оси диафиза по отношению к перпендикуляру, восстановленному от горизонтальной плоскости мыщелков |
|
СКД |
Степень изогнутости диафиза или расстояние между передней поверхностью диафиза на середине длины кости и плоскостью, проведенной между задними поверхностями головки и мыщелков, измеренное в сагиттальной плоскости |
Таблица 3
Остеометрия структур дистального эпифиза
|
Наименование структуры |
Расшифровка измерений |
|
ШДЭ |
Ширина дистального эпифиза – наибольшее расстояние между надмыщелками |
|
ШММЯ |
Ширина межмыщелковой ямки – внутреннее расстояние между краями ямки |
|
ШНК |
Ширина суставной поверхности для надколенника |
|
ØММ |
Сагиттальный размер медиального мыщелка |
|
ШММ |
Ширина суставной поверхности медиального мыщелка для бедренных костей |
|
ШЛМ |
Ширина суставной поверхности латерального мыщелка для бедренных костей |
|
ØЛМ |
Сагиттальный диаметр латерального мыщелка |
|
ШМНМ |
Ширина медиального надмыщелка – расстояние, измеренное горизонтально от максимально выступающей точки надмыщелка до края суставной поверхности |
|
ШЛНМ |
Ширина латерального надмыщелка – расстояние, измеренное горизонтально от максимально выступающей точки надмыщелка до края суставной поверхности |
Все цифровые значения размеров структур, измеряемые в миллиметрах, на следующем этапе делились на величину поперечного диаметра диафиза кости, измеренного на середине длины кости, что позволило учесть влияние соматотипа людей, веса и степени развития костяка у животных на формирование структурной организации кости. Дальнейшей обработке подвергались относительные значения изучаемых структур. Математические модели структурной организации костей были созданы по результатам многомерного корреляционного анализа методом Maximum Likelihood Factor с вращением Equamax normalized в демо-версиях программы Statistica 6.0–10.0. Сила коррелированности между структурами кости, составляющими один фактор при уровне статистической значимости 99,95 % (р≥ 0,05), составила |0,7–1,0|. Формирование уровней структурной организации костей шло путем уменьшения доли выделенной дисперсии в каждом последующем факторе. Все требования к выборкам для проведения факторного анализа были выполнены. В качестве корреляционной матрицы использовалась матрица корреляции Спирмена.
Результаты исследования и их обсуждение
Структурная организация бедренных костей независимо от видовой принадлежности оказалась асимметричной. Эта асимметрия проявилась различием как числа уровней структурной организации, так и параметров вовлеченных в организацию уровней. Так, правая бедренная кость человека (ПБК) имела четырехуровневую структуру, левая (ЛБК) –– трехуровневую. Структурная организация правой бедренной кости быка имела 3 уровня, левой – 4. Структура костей галопирующих животных – собак и кроликов – показала 4 уровня. Модели структурной организации бедренных костей, созданные по результатам факторного анализа, представлены на рисунках 1–4.
В структуре ПБК быков выделено 14 коррелирующих параметров, ЛБК – 15. У собак справа – 12, слева – 15; у кроликов справа – 24, слева – 20. У человека независимо от принадлежности к стороне – 19 (табл. 4).
Таблица 4
Межвидовое распределение числа коррелирующих параметров в структуре бедренной кости
|
вид |
Bos taurus taurus, L.1758 |
Canis lupus familiaris, L.1758 |
Oryctolagus cuniculus, L.1758 |
Современный человек |
||||
|
Часть кости |
ПБК |
ЛБК |
ПБК |
ЛБК |
ПБК |
ЛБК |
ПБК |
ЛБК |
|
Проксимальный эпифиз |
6 |
4 |
5 |
6 |
12 |
10 |
9 |
6 |
|
Диафиз |
3 |
5 |
2 |
4 |
5 |
1 |
3 |
4 |
|
Дистальный эпифиз |
5 |
6 |
5 |
5 |
7 |
9 |
7 |
9 |
|
Всего |
14 |
15 |
12 |
15 |
24 |
20 |
19 |
19 |
Примечание: ПБК – правая бедренная кость, ЛБК – левая бедренная кость.
У каждого из изучаемых видов были выделены системно стабильные параметры, присутствующие в структурной организации бедренных костей обеих конечностей представителей одного вида животных (рис. 1–4). Таких базовых параметров больше всего выделено у человека и кроликов – 14. В структуре бедренных костей быков данных параметров выявлено 6, собак – 4 (рис. 1–4). Параметры, присутствующие в структурной организации бедренных костей одной из конечностей, были определены как нестабильные.
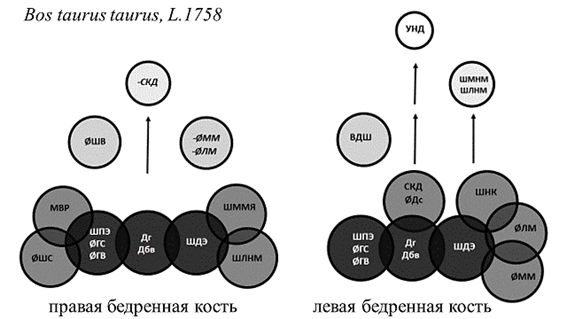
Рис. 1. Структурная организация бедренной кости быков
Примечание: Темно-серым цветом обозначены стабильные параметры, влияющие на структуру обеих костей. Курсивным шрифтом со знаком «минус» обозначены параметры, оказывающие отрицательное влияние.
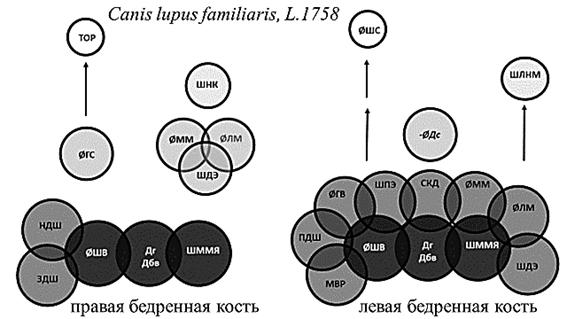
Рис. 2. Структурная организация бедренной кости собак
Примечание: Темно-серым цветом обозначены стабильные параметры, влияющие на структуру обеих костей. Курсивным шрифтом со знаком «минус» обозначены параметры, оказывающие отрицательное влияние.
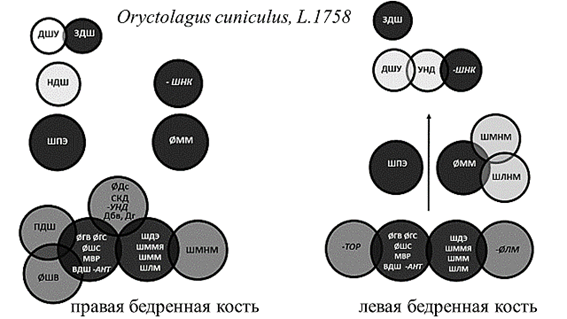
Рис. 3. Структурная организация бедренной кости кроликов
Примечание: Темно-серым цветом обозначены стабильные параметры, влияющие на структуру обеих костей. Курсивным шрифтом со знаком «минус» обозначены параметры, оказывающие отрицательное влияние.
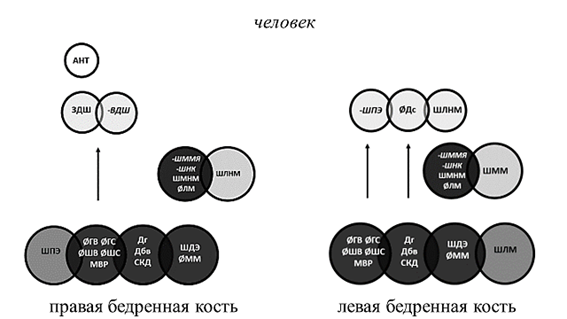
Рис. 4. Структурная организация бедренной кости человека
Примечание: Темно-серым цветом обозначены стабильные параметры, влияющие на структуру обеих костей. Курсивным шрифтом со знаком «минус» обозначены параметры, оказывающие отрицательное влияние.
У человека, собак и быков общим стабильным структурообразующим параметром первого уровня явилась длина кости, измеренная как от головки, так и от большого вертела. Базовая структура ЛБК кроликов лишена значимых корреляционных нагрузок на данный параметр, что может быть связано с асимметричностью конечностей в реализации опорной функции. Независимо от принадлежности к виду и стороне тела в основе структурной организации бедренной кости лежат параметры, отвечающие за передачу веса по оси конечности и осуществление сгибательно-разгибательных движений в обоих суставах. Эти параметры присутствуют у большинства исследуемых видов на костях обеих сторон тела. Это размеры головки, шейки, ширина дистального эпифиза и сагиттальный диаметр медиального мыщелка (рис. 1, 3, 4).
С расширением амплитуды сгибательно-разгибательных движений в тазобедренном суставе у животных, способных к практически вертикальному выталкиванию тела при прыжках [5] (кролик), и у человека в группе стабильных системообразующих параметров первого уровня появляются факторные нагрузки на межвертельное расстояние (рис. 3, 4). Изменения структурной организации проксимального эпифиза кроликов, связанные с прыжками, проявились факторными нагрузками на параметр, не только отражающий стереометрию проксимального эпифиза и гармоничность биомеханики животного, но и обеспечивающий передачу веса от центра тяжести на задние конечности. Это задняя длина шейки бедренной кости и величина антерверсии, одинаково вовлеченные в структурную организацию контрлатеральных костей у животных с симметричным типом локомоции – кроликов (рис. 3).
Выделение медиального мыщелка как стабильного структурообразующего параметра указывает на передачу силовых нагрузок через медиальный мыщелок независимо от видовой принадлежности и латерализации конечности. Отрицательные корреляционные нагрузки на ширину суставной поверхности для надколенника, характерные для человека и кроликов, указывают на увеличение роли данного параметра при переходе к прямохождению. У животных надколенник расположен значительно выше плоскости коленного сустава и играет в большей степени роль блока, увеличивающего силу четырехглавой мышцы бедра [6, 7]. У человека надколенник расположен несколько ниже и не только увеличивает силу четырехглавой мышцы бедра, но и ограничивает амплитуду разгибания в коленном суставе, что препятствует опрокидыванию корпуса при ходьбе [8]. Изменения структурной организации дистального эпифиза кости при переходе к прямохождению характеризуются отрицательными корреляционными нагрузками на ширину межмыщелковой ямки бедренной кости человека (рис. 4). Данный параметр определяет длину крестообразных связок и степень фиксации менисков. У животных, не имеющих механизма «запирания» коленного сустава при трансляции веса по оси конечности [6, 9, 10], – собак и кроликов – нагрузки на данный параметр будут положительными. У быков, имеющих фиброзный тяж, фиксирующий коленный сустав при опоре, факторные нагрузки неинформативны (рис. 1–3).
Системно нестабильные параметры, присутствующие только на одной из костей, вызывают максимальный интерес, поскольку отражают изменения структурной организации бедренной кости, связанные с особенностями локомоции. Асимметричность вовлечения параметров эпифизов кости в структурную организацию указывает, на наш взгляд, на морфофункциональную дифференциацию контрлатеральных костей на уровне эпифизов, в большей степени характерную для человека и животных с асимметричным способом локомоции (рис. 1–4).
Так, структурная организация проксимального эпифиза ПБК человека отличается положительными корреляционными нагрузками на ширину проксимального эпифиза, заднюю длину шейки и угол антеверсии, отрицательными нагрузками на верхнюю длину шейки, слева – отрицательными нагрузками на ширину проксимального эпифиза. У быков, которые, как и человек, являются представителями животного мира с асимметричной локомоцией, в структуре проксимального эпифиза ПБК присутствуют нагрузки на шейку и межвертельное расстояние, слева только на верхнюю длину шейки, что также указывает на морфофункциональную асимметрию. У животных, способных к одновременному отрыву задних лап при типичной локомоции, асимметричность организации проксимального эпифиза выражена меньше.
Асимметричность организации дистального эпифиза выражена значительно меньше, чем проксимального эпифиза, и проявляется миграцией параметров между уровнями структурной организации. При этом у быков в структурной организации дистального эпифиза ЛБК присутствуют нагрузки на ширину медиального надмыщелка. Степень выраженности медиального надмыщелка зависит от активности приводящих мышц, обеспечивающих постуральную устойчивость животного [5, 9]. Дистальный эпифиз ЛБК кроликов и собак в своей структурной организации имеет нагрузки на ширину латерального надмыщелка. У человека ширина латерального надмыщелка влияет на структурную организацию ПБК. От латерального надмыщелка у человека и животных начинаются латеральное брюшко икроножной мышцы и подошвенная мышца. Они обеспечивают не только сгибание в коленном суставе, но и пронацию голени. Нестабильные параметры дистального эпифиза указывают на асимметричность реализации двигательной функции как на уровне коленного сустава, так и нижележащих отделов конечности.
Заключение
Таким образом, структурная организация бедренной кости зависит от функциональной активности конечности. Увеличение последней приводит к росту числа стабильных структурообразующих параметров.
Для структурной организации бедренной кости человека и животных характерна латентная морфофункциональная асимметрия, свидетельствующая о различиях в функционировании эпифизов контрлатеральных костей.
Структурная организация бедренной кости человека подверглась большой перестройке, связанной с прямохождением. Она отличается увеличением числа коррелирующих параметров проксимального эпифиза при сохранении уровня коррелированности параметров дистального эпифиза.
Библиографическая ссылка
Яшина И.Н., Клочкова С.В., Иванов А.В. ИЗМЕНЕНИЯ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕДРЕННОЙ КОСТИ В РЯДУ «БЫК – СОБАКА – КРОЛИК – ЧЕЛОВЕК» // Современные проблемы науки и образования. 2019. № 2. ;URL: https://science-education.ru/ru/article/view?id=28633 (дата обращения: 02.08.2026).
DOI: https://doi.org/10.17513/spno.28633