



Нейротрансплантация рассматривается как перспективный подход для восстановления поврежденной нервной системы. Мозг является иммунопривилегированной областью организма, поэтому трансплантированная в него нервная ткань, состоящая из прогениторных и фетальных клеток, не отторгается и продолжает дифференцировку. В условиях нейротрансплантации создаются особые условия для развития аксональных путей в мозге: незрелые клеточные предшественники развиваются в микроокружении зрелого мозга и встраиваются в уже существующие нейрональные системы. Возможность формирования синаптических связей между нейронами трансплантированной ткани и мозгом реципиента определяется способностью нервной ткани разного возраста к морфофункциональным пластическим преобразованиям и к взаимной адаптации. Известно, что при трансплантации могут устанавливаться как типичные, так и атипичные, не свойственные контактирующим отделам мозга синаптические связи [1; 2].
При многих неврологических заболеваниях значительные нарушения происходят в гиппокамповой формации. Эпилепсия, например, приводит к значительной гибели нейронов, аберрантному спрутированию аксонов гранулярных нейронов зубчатой фасции (мшистых волокон гиппокампа) и дальнейшему распространению патологической активности в мозге. Мшистые волокна заканчиваются уникальными гигантскими синаптическими окончаниями на апикальных дендритах пирамидных нейронов поля СА3 гиппокампа. Ранее мы показали, что между трансплантатами зубчатой фасции гиппокампа и неокортексом реципиента формируются функциональные синаптические контакты, хотя в норме эти два отдела мозга не взаимодействуют. При этом в пресинаптических терминалях таких синапсов серьезно изменяется везикулярно-химический состав: уменьшается количество везикул, содержащих глутаматергический трансмиттер, и увеличивается число нейропептидных гранул [1; 3]. В то же время ничего не известно о структурной перестройке в нейронах мозга реципиента, которые взаимодействуют с чужеродными аксонами, проникающими из трансплантата. Целью настоящей работы являлось изучение структурно-химической реорганизации в постсинаптических компартментах таких неспецифических контактов. Для этого проводили сравнение ультраструктуры гигантских синапсов мшистых волокон гиппокамповой формации крыс, образованных в неокортексе после трансплантации зубчатой фасции, и таковых в здоровом мозге.
Материалы и методы исследования
Эксперименты проводили на крысах породы Вистар с соблюдением рекомендаций к работе с животными (ГОСТ РИСО10993-2-2009). Все процедуры проводили под нембуталовым наркозом (40 мг/кг) и местной анестезией. Донорским материалом для аллотрансплантации в соматосенсорную область неокортекса служили закладки зубчатой фасции 20-дн. плодов; в качестве реципиентов использовали пять половозрелых крыс-самцов той же породы. Через 5 месяцев после операции мозг фиксировали 2.5% раствором глутарового альдегида, выделяли область неокортекса с трансплантатом и нативный гиппокамп из другого полушария мозга для контроля. Далее материал дофиксировали четырехокисью осмия и обрабатывали для электронной микроскопии по стандартной методике. Синаптические окончания мшистых волокон идентифицировали по большим размерам терминалей (не менее 3-4 мкм в диаметре) и другим детерминантным признакам, известным из литературы и собственных наблюдений [1; 3; 4]. Для количественного анализа использовали по 50 случайно выбранных микроизображений гигантских синапсов (n = 50) из экспериментальных и контрольных образцов с хорошо выраженными активными зонами на дендритных шипиках и адгезивными соединениями на стволах дендритов. Электронно-микроскопические фотоизображения сканировали, сохраняли в виде компьютерных файлов и производили морфометрический анализ с помощью программы UTHSCSA Image Tool. Сравнительный анализ проводили по следующим показателям: число адгезивных контактов между аксональной терминалью и дендритом и их суммарная длина (в мкм) в среднем на синапс, а также отношение суммарной длины адгезивных соединений к полной протяженности зоны прилегания терминали к поверхности дендрита, в процентах. Статистическую обработку результатов проводили с помощью пакета программ Origin 6.0. Вычисляли средние значения для групп показателей (М) и стандартные ошибки (m). Для определения уровней значимости различий (Р) каждого показателя использовали параметрический t-критерий Стьюдента.
Результаты исследования и их обсуждение
При электронно-микроскопическом изучении контрольного материала (прозрачного слоя поля СА3 нативного гиппокампа) легко идентифицировались гигантские синаптические окончания аксонов гранулярных нейронов зубчатой фасции. Они компактно располагались между крупными профилями апикальных дендритов пирамидных нейронов и соответствовали известным морфологическим критериям [4]. Кроме гигантских размеров аксональных терминалей (до 5-6 мкм в диаметре), от обычных синапсов их отличало наличие двух типов функциональных контактов: синаптические активные зоны с дендритными шипиками и симметричные адгезионные соединения с поверхностью самих дендритов. При удачном плане среза можно было наблюдать места отрастания разветвленных шипиков от дендритных стволов (рисунок 1).
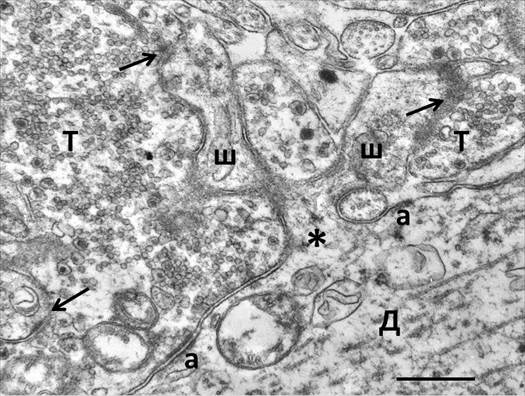
Рис. 1. Гигантское синаптическое окончание мшистого волокна в гиппокампе крысы.
Д – дендрит; Т – синаптическая терминаль; а – адгезивное соединение; ш – головка разветвленного шипика; стрелка – синаптический контакт; звездочка – место ответвления шипика от дендрита. Масштаб: 0.5 мкм
Их синаптические терминали были плотно заполнены малыми светлыми и большими гранулярными синаптическими пузырьками, среди которых наблюдались также митохондрии и профили перерезанных дендритных шипиков с активными зонами, имеющими хорошо выраженные постсинаптические уплотнения. В цитоплазме головок разветвленных дендритных шипиков среди типичного для них филаментозного материала наблюдались вакуоли и цистерны эндоплазматического ретикулума, некоторые из которых были упорядочены в виде типичного шипикового аппарата. Кроме того, в дендритных шипиках иногда присутствовали свободные или сгруппированные рибосомы. Каждая гигантская синаптическая терминаль широко распластывалась по поверхности дендрита. Между смежными аксональной и дендритной мембранами наблюдались адгезивные соединения, около которых со стороны аксональной терминали никогда не было синаптических везикул. Из-за симметрично расположенного осмиофильного адгезивного материала с обеих сторон противостоящих мембран такие соединения называли десмосомоподобными, или puncta adherentia, контактами и рассматривали как места механического удержания сложного синаптического комплекса [4]. С помощью современных методов молекулярной биологии определен нейрохимический состав адгезивных соединений, который свидетельствует об их более значительной роли в координации морфофункциональных процессов в гигантских синапсах гиппокампа [5-7].
Для изучения структурной реорганизации синаптических окончаний мшистых волокон при контакте с чужеродными нейрональными мишенями был исследован прилежащий к трансплантату неокортекс мозга реципиента на глубину от 200 до 500 мкм. Гигантские синаптические терминали в неокортексе не располагались плотным слоем, как это типично для их места расположения в гиппокампе, а были распределены среди элементов соматосенсорной области достаточно рассеянно. В то же время вокруг некоторых дендритов наблюдались их скопления, которые, по-видимому, представляли собой последовательные синаптические бутоны одного и того же аксона. Такие синапсы являются химерами, пресинаптические отделы которых принадлежат нейронам трансплантатов, а постсинаптические – нейронам мозга реципиента. Принцип организации химерных синапсов мшистыми волокнами во многом совпадал с таковым в мозге in situ, т.е. интратерминальные химические синапсы устанавливались с инвагинированными дендритными шипиками, а адгезивные несинаптические соединения - с поверхностью дендритов. Примечательно, что дендритные шипики неокортекса, в норме имеющие классическую грибовидную форму, при контакте с мшистыми волокнами, проникающими из трансплантатов, часто приобретали разветвленные головки неопределенной конфигурации (рисунок 2).
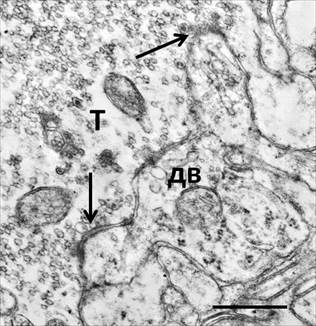
Рис. 2. Поперечный срез разветвленного дендритного выроста (дв), устанавливающего синаптические контакты с синаптической терминалью мшистого волокна трансплантированного нейрона в неокортексе реципиента
Обозначения те же, что на рис. 1.
Присутствие аномально разветвленных типов дендритных шипиков на дендритах неокортикальных нейронов свидетельствуют о том, что аксоны, врастающие из трансплантатов, индуцируют в мозге реципиента формирование соответствующих им сложных постсинаптических микровыростов. При этом шиповидные выросты в эктопических гигантских синапсах отличались от таковых в норме более богатым и разнообразным содержимым. Во многих из них встречались митохондрии, а рибосомы были собраны в полисомы или ассоциированы с цистернами гранулярного эндоплазматического ретикулума. Известно, что появление в дендритных шипиках органелл, ответственных за синтез белка, характерно для периодов пластической перестройки синаптического аппарата во время развития или функциональной активации нейронов [8]. Многообразие органелл внутри дендритных микровыростов в нашем экспериментальном материале, когда мшистые волокна формируют аберрантные функциональные связи, по-видимому, отражает процесс нейрохимической реорганизации в связи с адаптацией постсинаптических локусов нейронов неокортекса к проникающим из нейротрансплантата аксонам.
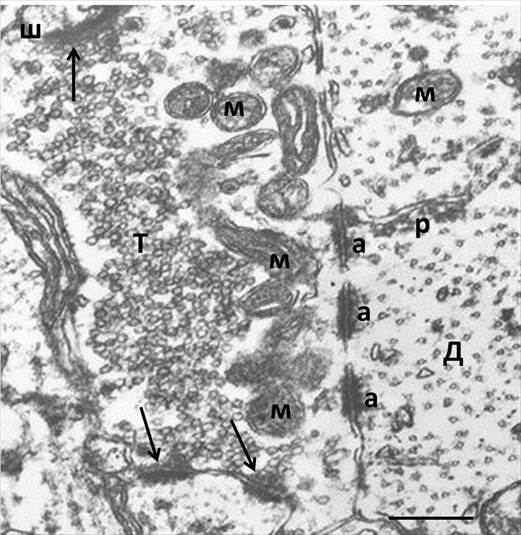
Рис. 3. Адгезивные соединения между дендритом и аксональной терминалью в эктопическом гигантском синаптическом окончании, сформированном в неокортексе реципиента после трансплантации
Обозначения: м – митохондрии; р – цистерны эндоплазматического ретикулума; другие обозначения те же, что на рис. 1.
Важные изменения ультраструктуры эктопических гигантских синапсов были обнаружены в топографии и выраженности адгезивных соединений, образованных между аксональными терминалями и дендритными стволами (рисунок 3). В первую очередь следует отметить, что симметрично адгезивным участкам в аксональной терминали, принадлежащей трансплантату, со стороны дендритного ствола, принадлежащего нейронам реципиента, также присутствовали аналогичные осмиофильные уплотнения, хотя их наличие не свойственно неокортикальным дендритам в норме. Это придавало внесинаптическим адгезивным зонам в эктопических окончаниях мшистых волокон вид соединений типа puncta adherentia, характерных для гиппокамповой формации. Межклеточное пространство в участках внесинаптических адгезионных соединений было заполнено волокнистым материалом, который иногда был организован в виде поперечных мостиков, демонстрирующих бинаправленные межклеточные взаимодействия молекул клеточной адгезии через такие зоны. Иногда участки адгезии занимали основную часть области аппозиции терминали к дендритному стволу или концентрировались в местах ответвления дендритных шипиков. В некоторых синапсах они распространялись на ножки дендритных шипиков и сливались с активными зонами, хотя в норме эти два типа контактов всегда топографически разнесены. Известно, что при дифференцировке гигантских синапсов в онтогенезе нарождающиеся примитивные активные зоны расположены на дендритах и колокализованы с адгезивными контактами [9]. Эти литературные данные позволяют нам предположить, что и в условиях гетеротопической трансплантации зубчатой фасции, когда формируются аберрантные синаптические связи, соединения puncta adherentia также принимают участие в генезе новых аксо-шипиковых активных зон.
При микроскопическом анализе аксо- и дендроплазмы вблизи адгезионных соединений в эктопических гигантских синапсах неокортекса были выявлены признаки повышенной функциональной активности по сравнению с идентичными локусами в гигантских синапсах in situ. Так, со стороны пресинаптического бутона около них скапливались митохондрии, что свидетельствует о б?льшей энергозависимости процесса аберрантной коммуникации через адгезивные специализации. Со стороны ствола дендрита наблюдалась концентрация органелл, имеющих прямое отношение к синтезу метаболитов. Среди них были идентифицированы свободные и прикрепленные к цистернам шероховатого эндоплазматического ретикулума рибосомы, митохондрии и другие микроструктуры. Некоторые группы канальцев эндоплазматического ретикулума находились в непосредственном контакте с конгломератами адгезивного материала (рисунок 3).
Проведенный количественный анализ подтвердил наблюдения об усилении адгезии вне активных зон эктопических синапсов (таблица). По сравнению с контролем в них обнаружено достоверное увеличение количества дискретных адгезионных соединений и их суммарной протяженности более чем в 2 раза. В среднем количество дискретных участков в синапсах, сформированных в неокортексе после трансплантации, было 3.7±0.4, а в контрольных синапсах только 1.7±0.3. Процент, который занимала суммарная длина отдельных участков зон адгезии от полной длины прилегания аксональной терминали к дендриту в эктопических синаптических окончаниях, в среднем также значительно превышал их значения в контрольных синапсах (29.0±7.0% против 8.0 ±3.0%).
Количественный анализ адгезионных соединений (АС) в гигантских синапсах мшистых волокон в контроле и после трансплантации. Средние значения и стандартные ошибки
(M ± m); Р – уровень различий показателей для каждого параметра
|
Параметры |
Контроль |
Трансплантация |
Р |
|
Среднее число АС в синапсе |
1.7 ± 0.3 |
3.7 ± 0.4 |
0.001 |
|
Суммарная длина АС в синапсе (мкм) |
0.31 ± 0.08 |
0.71 ± 0.12 |
0.009 |
|
Суммарная длина АС/полная длина аппозиции терминали к дендриту (%) |
8.0 ± 3.0 |
29.0 ± 7.0 |
0.009 |
Существенное значение внесинаптических адгезионных соединений в формировании эктопических синапсов в условиях нейротрансплантации подтверждают известные литературные данные об их нейрохимическом составе. В основу молекулярной организации этих контактов входят кадгерин-катениновая и нектин-афадиновая адгезивные системы, а также синаптическая адгезионная молекула S-SCAM [5-7]. Классические кадгерины играют ключевые роли в развитии и физиологии нервной системы. Они участвуют в морфогенезе дендритных шипиков, прикрепляя AMPA рецепторы к активной зоне через белки катенины [6; 10]. Нектины, располагаясь на обеих сторонах puncta adherentia, взаимодействуя через афадин с актиновым цитоскелетом, также вовлекаются в моделирование синапсов [11].
Заключение
Электронно-микроскопическое сравнение гигантских синапсов мшистых волокон, сформированных в неокортексе реципиента после трансплантации зубчатой фасции крыс, с таковыми в гиппокампе in situ показало, что под влиянием аксонов, прорастающих из нейротрансплантатов, в постсинаптических компартментах эктопических синапсов происходила значительная структурная реорганизация. Она касалась основных детерминантных признаков гигантских синапсов. На дендритах неокортекса, контактирующих с аксонами трансплантированных нейронов, были обнаружены несвойственные им в норме разветвленные дендритные шипики, инвагинирующие аксональные терминали для формирования интратерминальных синаптических активных зон. В местах прилегания крупных дендритных ветвей к гигантским терминалям, в примембранных локусах дендритов неокортекса были идентифицированы скопления осмиофильного материала, наподобие того, который присутствует в гиппокампе in situ на местах адгезивных соединений типа puncta adherentia. Количественный морфометрический анализ показал увеличение протяженности таких адгезионных соединений в эктопических гигантских синапсах более чем в 2 раза. Важно, что эти морфологические изменения в эктопических синапсах сопровождались появлением в цитоплазме дендритов и дендритных шипиков дополнительных органелл (полисом, цистерн гранулярного эндоплазматического ретикулума, митохондрий), имеющих отношение к процессам клеточного синтеза. Полученные данные свидетельствуют о том, что в условиях трансплантации нейроны мозга реципиента не только перестраивают свою микроструктуру, но и адаптируют метаболический аппарат к установлению функциональных контактов с чужеродными аксонами, проникающими из нейротрансплантатов.
Работа выполнена при финансовой поддержке РФФИ (грант № 17-04-00786).
Библиографическая ссылка
Журавлева З.Н., Журавлев Г.И., Шубина Л.В. СТРУКТУРНАЯ РЕОРГАНИЗАЦИЯ НЕЙРОНОВ МОЗГА РЕЦИПИЕНТА ПРИ УСТАНОВЛЕНИИ СИНАПТИЧЕСКИХ КОНТАКТОВ С НЕЙРОТРАНСПЛАНТАТОМ // Современные проблемы науки и образования. 2018. № 6. ;URL: https://science-education.ru/ru/article/view?id=28356 (дата обращения: 02.07.2026).