



Известно, что особенностью физиологических систем у человека на Севере и в Арктике является функциональное напряжение [1-3]. Кроме того, в течение годовых циклов происходят адаптивные изменения практически во всех органа и системах. Так, например, под влиянием изменений естественной освещенности меняется функциональное состояние центральной нервной системы [4]. Перестройки различных функциональных систем, происходящие под влиянием суровых климатогеографических условий высоких широт, формируют новое состояние организма – экологическую адаптированность, которое достигается ценой определенной биосоциальной платы [5; 6]. В то же время активизация деятельности человека в Арктике и реализация новой стратегии социально-экономического развития северных регионов диктует необходимость изучения не только физиологических механизмов адаптационных перестроек, но и физиологических характеристик функциональных систем как результата этих перестроек. Формирование экологической адаптированности церебрального энергообмена происходит при значительном увеличении его интенсивности в центральной нервной системе, что соответствует признакам синдрома адаптационного профицита церебрального энергообмена [7].
Однако данных о формировании экологической адаптированности энергетического состояния головного мозга в климатогеографических условиях Арктической зоны явно недостаточно, не описаны ее структура и характер. Нет данных о влиянии различных факторов на особенности формирования экологической адаптированности, как то: возраста, пола, состояния здоровья, межполушарной асимметрии головного мозга, исходного вегетативного тонуса и др., что, по нашему мнению, является весьма важным для разработки вопросов сохранения и укрепления здоровья, обоснования и применения лечебно-диагностических и коррекционно-реабилитационных мероприятий.
Цель настоящей работы – определить особенности развития и состояния экологической адаптированности церебрального энергообмена у молодых жителей Арктического региона при разном исходном вегетативном тонусе.
Материал и методы исследования. На первом этапе данной работы определялся исходный вегетативный тонус методом кардиоритмографии с регистрацией фоновой электрокардиограммы (ЭКГ) в состоянии относительного покоя (после предварительно отдыха в положении сидя) в первой половине дня с помощью аппаратно-программного комплекса «Варикард». Исследовались временные параметры вариабельности сердечного ритма у 103 молодых людей (45 юношей, 58 девушек, средний возраст 19±0,22 года), родившихся и постоянно проживающих в г. Архангельске: стандартное отклонение средней продолжительности интервалов RR (SDNN); квадратный корень из среднего квадрата разности последовательных интервалов RR (RMSD); процент последовательных интервалов RR, разница между которыми превышает 50 мс (pNN50, %). Увеличение временных параметров вариабельности сердечного ритма расценивали как усиление парасимпатических влияний (группа ваготоников), снижение – как активацию симпатических влияний (группа симпатотоников). Оценивали параметры сердечного ритма в трех частотных диапазонах: высокочастотные колебания (0,4–0,15 Гц), HF (мс2), характеризующие влияние парасимпатического отдела вегетативной нервной системы (ВНС); низкочастотные колебания (0,15–0,04 Гц), LF (мс2), отражающие преимущественно активность симпатического отдела; сверхнизкочастотные колебания (0,04–0,003Гц), VLF (мс2), которые характеризуют степень связи автономных сегментарных уровней регуляции кровообращения с надсегментарными. Также оценивали суммарную мощность спектра TF (мс2) и рассчитывали индекс LF/HF, отражающий баланс симпатической и парасимпатической активности [8].
В дальнейшем исследование церебральных энергетических процессов проводилось у двух групп: 1 группа – 30 человек с симпатотонией и 2 группа – 21 человек с ваготонией. Поскольку показатели у юношей и девушек не имели достоверных отличий, то они были объединены в единые группы. К группе со смешанным вегетативным тонусом (амфотония) были отнесены 52 человека (51%).
Для оценки церебральных энергетических процессов осуществлялась регистрация уровня постоянного потенциала (УПП) головного мозга на аппаратно-программном комплексе «Нейро-КМ». Распределение УПП регистрировали монополярно с помощью хлорсеребряных электродов с контролируемой разностью потенциалов 1 мВ и усилителя постоянного тока с входным сопротивлением 100 кОм. Референтный электрод устанавливался на запястье руки. Активные электроды располагали на голове в лобной, центральной, затылочной областях вдоль сагиттальной линии и парасагиттальной в правом и левом височных отделах по схеме 10–20. Регистрацию осуществляли через 5 минут после наложения электродов с прокладками, смоченными насыщенным раствором NaCl для снижения кожного сопротивления и уменьшения величины кожных потенциалов. Во время регистрации проводили контроль кожного сопротивления.
Статистический анализ полученных результатов проводили с использованием программных продуктов SPSS-20 фирмы IBM for Windows. Проверка на нормальность распределения осуществлялась тестом Колмогорова-Смирнова. В случае нормального распределения переменных применялись параметрические методы для независимых выборок (t-Стьюдента). Результаты параметрических методов обработки данных представлялись в виде среднего значения (M) и стандартного отклонения (σ). Для всех приведенных результатов различия считались значимыми при уровне p<0,05. Особенности структуры и взаимоотношений показателей экологической адаптированности церебрального энергетического метаболизма у молодых людей устанавливались с помощью факторного анализа с варимакс-вращением.
Методика проведенного научного исследования составлена и проведена в соответствии со статьями 5, 6 и 7 Всеобщей декларации о биоэтике и правах человека.
До включения в исследование у всех участников было получено письменное информированное согласие в соответствии с принципами Хельсинкской декларации. При сборе материала для составления выборок соблюдались все необходимые условия: регистрация физиологических показателей проводилась в утреннее время, через 1,5–2 ч после приема пищи при максимальном физическом и психическом покое.
Результаты исследования и обсуждение. Сравнительный анализ значений распределения УПП выявил некоторые особенности энергозатрат по коре головного мозга у жителей Арктического региона в зависимости от вегетативного тонуса (таблица).
Распределение УПП головного мозга в однополярных отведениях у молодых людей с различным вегетативным тонусом, проживающих в Арктическом регионе (M±σ), мВ
|
Показатель |
Симпатотоники (n = 30) |
Парасимпатотоники (n = 21) |
|
Fz |
14,28±2,28 |
10,58±2,02 * |
|
Cz |
17,23±1,94 |
17,82±2,30 |
|
Oz |
13,16±2,25 |
15,86±2,06 |
|
Td |
12,04±2,12 |
9,37±1,70 * |
|
Ts |
10,78±2,12 |
9,49±1,36 * |
|
Sum |
67,49±9,27 |
63,12±7,61 |
Примечание. Звездочкой * обозначены статистически значимые различия между группами, p<0,05.
Так, у молодых людей симпатотоников значения УПП во всех пяти монополярных отведениях были выше нормативных данных средней полосы. Минимальное отклонение от эталона регистрировалось в левом височном отделе. А в группе ваготоников значения уровня потенциала в височных отведениях были ниже нормативов (рис. 1), что указывает на минимизацию энергетических процессов в данных отделах мозга.
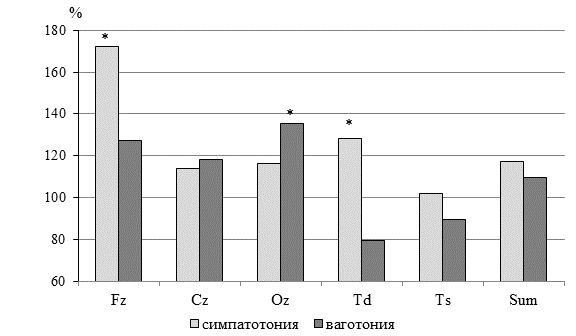
Рис. 1. Распределение УПП головного мозга у молодых людей Арктической зоны РФ в %% по отношению к нормативным значениям средней полосы
Примечание. Звездочкой * обозначены статистически значимые различия между фактическими и нормативными показателями, p<0,05.
Межгрупповое сравнение свидетельствует о преобладании энергетических процессов во фронтальных отделах мозга у симпатотоников, что находит свое отражение в высоких значениях УПП в лобном отведении и положительных значениях локального (Fz-X) и межэлектродных градиентов постоянных потенциалов.
В группе ваготоников повышенная интенсивность энергетических процессов регистрируется в затылочном отведении при резком падении уровня потенциала в височных отделах и снижение энергозатрат во фронтальной области.
Обращает на себя внимание и значения межвисочной разности потенциала, характеризующие межполушарную асимметрию головного мозга [9]. В группе симпатотоников разность Td–Ts=1,25 мВ, что указывает на доминирование активности правого полушария. В группе ваготоников данная разность отрицательна Td–Ts=-0,12 мВ и имеет значение меньше одного милливольта, что может свидетельствовать о тесном межполушарном взаимодействии с незначительным преобладанием активности левого полушария.
Суммарные значения УПП в группе симпатотоников были на 4,37% выше, чем в группе ваготоников. В группе ваготоников УПП в затылочных отделах был несколько выше, чем у симпатотоников.
Факторный анализ с использованием варимакс-ротации также выявил различия в состоянии экологической адаптированности церебральных энергетических процессов у молодых жителей Арктической зоны РФ при симпатотонии и ваготонии (рис. 2).
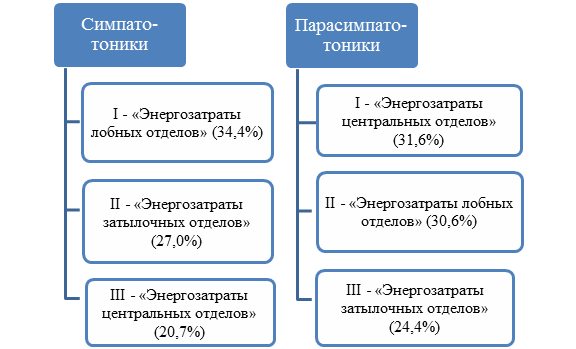
Рис. 2. Факторная структура экологической адаптированности церебрального энергообмена у молодых лиц с различным вегетативным тонусом
Факторные структуры, построенные по результатам исследования, указывают на различия энергетических процессов в коре головного мозга у симпатотоников и ваготоников. Так, первым фактором у симпатотоников являются «энергозатраты лобных отделов мозга». Вторым по значимости фактором выступают «энергозатраты затылочных отделов мозга», а третьим – «энергозатраты центральных отделов». У ваготоников первым является фактор «энергозатраты центральных отделов мозга», вторым – «энергозатраты лобных отделов», а третьим – «энергозатраты затылочных отделов». Следовательно, и распределение значений УПП, и факторные структуры указывают на различные механизмы церебрального энергетического обмена у молодых людей с разным типом регуляции ритма сердца. Так, у симпатотоников значения УПП выше по всей коре головного мозга в сравнении с парасимпатотониками. Известно, что благодаря работе симпатического отдела вегетативной нервной системы скорость окислительных процессов увеличивается [10]. Все это, видимо, и приводит к росту УПП в целом по всем отделам коры больших полушарий.
Кроме того, активность симпатического отдела ВНС отмечается при стрессовых ситуациях, с состоянием стресса также связывают повышение значений уровня постоянного потенциала мозга [11]. Высокие значения уровня постоянного потенциала в группе симпатотоников отмечаются в лобных отделах. Первый фактор в факторной структуре перераспределения энергетических затрат по коре также указывает на активизацию энергетических процессов лобного отдела и фронтальных структур головного мозга. Одной из главных функций лобных долей является анализ и синтез поступающей информации, формирование целевых установок и алгоритмизация дальнейшего функционирования организма.
В группе ваготоников, напротив, выявлено снижение значений УПП в сравнении с симпатотониками, а в височных отделах даже ниже нормативных. Данный факт, вероятно, связан с экономизацией вегетативных функций при повышении активности парасимпатического отдела ВНС. При этом увеличение энергозатрат регистрируется в затылочной области. Высокое значение первого фактора «энергозатраты центральных отделов коры головного мозга» указывает на тесное взаимодействие коры и подкорковых структур (лимбическая система, ретикулярная формация). Отсутствие явного проявления межполушарного доминирования свидетельствует о значительном количестве межполушарных связей, указывает на процесс адаптивных перестроек функциональных систем организма [12].
Заключение. Таким образом, факторные структуры экологической адаптированности церебрального энергообмена у симпатотоников и ваготоников свидетельствуют о том, что нейрофизиологические механизмы энергообеспечения в коре головного мозга у этих групп имеют различный характер: у симпатотоников протекают более напряженно, что находит свое отражение в повышенных значениях УПП во фронтальных и правовисочном отделах. Известно, что правое полушарие более автономно, чем левое, и менее подвержено влиянию корригирующих обратных связей. Кроме того, адаптационные перестройки, как правило, связывают с активацией правого полушария [12]. Высокие значения энергозатрат лобных отделов у симпатотоников, очевидно, свидетельствуют о значительном напряжении управляющих функций и высокой централизации регуляторных процессов [7]. То есть у симпатотоников адаптационные процессы еще не закончены и экологическая адаптированность энергетических процессов окончательно не сформирована.
У ваготоников энергозатраты в лобных отделах представлены во втором факторе, значимость их ниже, чем у симпатотоников. Нейрофизиологические процессы, связанные с энергообеспечением головного мозга, у ваготоников протекают менее напряженно. А высокие значения энергозатрат отмечаются в центральных отделах, которые тесно связаны с подкорковыми структурами головного мозга. То есть у ваготоников выявлено более тесное взаимодействие коркового и подкоркового уровней, что, в свою очередь, приводит к менее напряженному функционированию центральной нервной системы.
Исследование выполнено при финансовой поддержке РФФИ и правительства Архангельской области в рамках научного проекта № 18-44-290006.
Библиографическая ссылка
Волокитина Т.В., Аникина Н.Ю., Котцова О.Н., Грибанов А.В. ПРОЯВЛЕНИЯ ЭКОЛОГИЧЕСКОЙ АДАПТИРОВАННОСТИ ЦЕРЕБРАЛЬНОГО ЭНЕРГООБМЕНА У МОЛОДЫХ ЖИТЕЛЕЙ АРКТИЧЕСКОГО РЕГИОНА С РАЗЛИЧНЫМ ВЕГЕТАТИВНЫМ ТОНУСОМ // Современные проблемы науки и образования. 2018. № 6. ;URL: https://science-education.ru/ru/article/view?id=28335 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/spno.28335