



Участки акваторий рек, на которых расположены зимовальные ямы, всегда имели высокую ценность как в рыбохозяйственном отношении, так и для местного населения. На зимовальных ямах заготавливали в значительных количествах такие ценные виды рыб, как нельма, стерлядь, сибирский осетр. Значительные скопления рыб на таких участках рек отмечены не только в зимний период, но и практически круглогодично. При этом структура скоплений подвержена значительным изменениям как в количественном, так и в качественном (таксономическом, размерном) отношении. В связи с этим целью работы было установить таксономический состав (в том числе долю и численность ценных видов рыб – сиговых и осетровых) скоплений на русловой яме в бассейне р. Иртыш по окончании зимовки рыб (после распаления льда и активного подъема уровня воды) методом гидроакустической съемки.
Материал и методика. Гидроакустические исследования распределения рыб выполнены в весенний период (15 мая 2015 г) в акватории Кондинской русловой зимовальной ямы. Данный водный макробиотоп расположен на участке Нижнего Иртыша (90-91 км от устья) (60°42'28,21'' С.Ш., 69°40'34.88'' В.Д.). Исследуемый участок реки находится в пределах Ханты-Мансийского района Ханты-Мансийского автономного округа на месте впадения р. Конда в р. Иртыш.
В период проведения научно-исследовательских работ максимальные глубины в акватории ямы достигали 40 м и более, ширина русла р. Иртыш превышала 500 м. Площадь исследуемой акватории превышала 117 га.
Гидроакустическую съемку акватории выполняли с помощью программно-технического гидроакустического комплекса AsCor. Работа данного комплекса основана на использовании эхолота Furuno с вертикальным обзором (частоты 50 и 200 kHz). Эхосигнал эхолота подвергался обработке аналогово-цифровым преобразователем. Передвижение судна по акватории для выполнения гидроакустической съемки (рис. 1) осуществляли согласно общепринятым методикам [1]. Таксономическую принадлежность рыб, зарегистрированных гидроакустическим методом, в дальнейшем выполняли с помощью компьютерной программы Таxonomy в лабораторных условиях. Идентификация зарегистрированных групп рыб производилась по следующим группам: карповые, окуневые, сиговые и нераспознанные. В бассейне р. Иртыш к последней группе рыб, по данным исследовательского коллектива Э.С. Борисенко [4], отнесены рыбы семейства осетровых, налимовых и щуковых. Из-за вытянутой формы плавательного пузыря их определение по форме отраженного эхосигнала затруднительно.
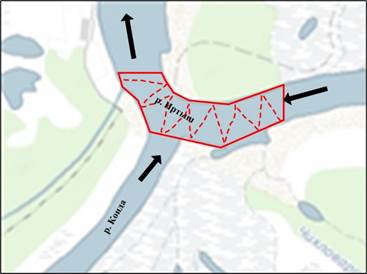
Рис. 1. Схема передвижения моторной лодки галсами (зигзагами) в акватории Кондинской русловой зимовальной ямы при проведении гидроакустической съемки (черными стрелками показано направление течения рек, красным цветом обозначены границы ямы, красным пунктиром обозначено движение судна)
Для контроля и определения видового состава рыбного населения выполняли контрольный лов рыбы разноячейными ставными и плавными сетями (размер ячеи 14, 25, 35, 45, 55, 65 мм, длина сети 35–75 м).
Результаты исследований и их обсуждение. В результате проведенного контрольного лова в акватории русловой ямы установлено, что рыбное население представлено следующими наиболее массовыми видами рыб: стерлядь (Acipenser ruthenus Linnaeus, 1758), сибирский осетр (Acipenser baerii Brandt), нельма (Stenodus leucichthys nelma Pallas, 1773), пелядь (Coregonus peled Gmelin, 1788), плотва (Rutilus rutilus Linnaeus, 1758), язь (Leuciscus idus (Linnaeus, 1758), елец (Leuciscus leuciscus (Linnaeus, 1758), лещ (Abramis brama (Linnaeus, 1758), золотой карась (Carassius carassius Linnaeus, 1758), серебряный карась (Carrassius auratus Linnaeus, 1758), окунь (Perca fluviatilis Linnaeus, 1758), ерш (Gimnocephalus cernuus Linnaeus, 1758), судак (Sander lucioperca Linnaeus, 1758), щука (Esox lucius Linnaeus, 1758), налим (Lota lota Linnaeus, 1758). Среди сиговых рыб доминирующим видом была нельма, ее доля в этой группе составила 97%, незначительное количество пеляди 2%. Среди группы нераспознанных рыб распределение рыб по семействам было следующим: осетровые 93%, налимовые 2,5%, щуковые 4,5%.
По данным гидроакустической съемки, общая доля группы карповых рыб составила 67,11%, окуневых 20,48%, сиговых 7,75%, нераспознанных 4,66%.
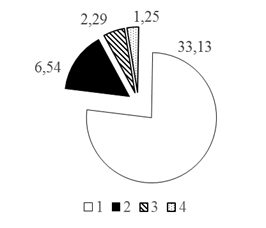
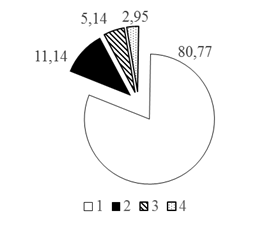
Для удобства анализа вертикального распределения рыб всю водную толщу условно разделили на 4 горизонта: 0-10, 10-20, 20-30 м и >30 м. В результате анализа вертикального распределения рыб установлено, что преобладающая часть рыбного населения - 81,45% от общего числа всех зарегистрированных рыб - распределена в двух первых горизонтах водной толщи 0-10 и 10-20 м. Доли рыб здесь составляли 43,21% и 38,24% соответственно (рис. 2 А, Б).
 А
А 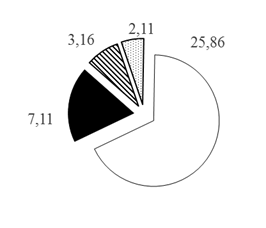 Б
Б
 В
В 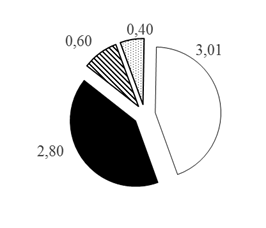 Г
Г
Рис. 2. Распределение различных таксономических групп рыб в акватории Кондинской русловой ямы по горизонтам глубин (А. 0-10 м; Б. 10-20 м; В. 20-30 м; Г.>30 м), в % от общего числа зарегистрированных рыб: 1. Карповые 2. Окуневые 3. Сиговые 4. Нераспознанные
В последующих горизонтах водной толщи Кондинской русловой ямы с глубинами 20-30 м и >30 м находилось значительно меньшее количество рыб, их процентное соотношение от общего числа зарегистрированных рыб составило 11,74% и 6,81% соответственно (рис. 2 В, Г). Такая закономерность уменьшения численности рыб естественна, поскольку русло любого водотока при поперечном рассмотрении уменьшается в направлении от поверхности воды до дна и имеет вид клина. Снижение численности рыб при уменьшении объема жизненного пространства закономерно, поскольку таким образом работают компенсаторные механизмы популяции, при разрежении плотности рост рыб активнее происходит рост рыб и снижается конкуренция за кормовые ресурсы [5].
Для карповых рыб отмечена следующая закономерность: происходит уменьшение количества рыб от поверхности ко дну в 11 раз с 33,13% в поверхностном горизонте 0-10 м до 3,01% в водной толще на глубинах >30 м (рис. 2А).
Для окуневых, сиговых и группы нераспознанных рыб вертикальное распределение в акватории водной толщи ямы происходит несколько другим образом: наибольшая их доля отмечена в горизонте 10-20 м: 7,11, 3,16, 2,11% соответственно, а уменьшение происходит сначала в направлении поверхностного горизонта водной толщи 0-10 м, затем в направлении придонного горизонта водной толщи с глубинами >30 м, где и отмечены минимальные значения их долей от общего числа зарегистрированных особей рыбного населения русловой ямы: 2,80, 0,60, 0,40% соответственно (рис. 2 Б-Г).
Разница максимального и минимального распределения окуневых и сиговых и нераспознанных групп рыб на глубинах 10-20 м и >30 м составляет распределения в 2,54, 5,27, 5,28 раза соответственно, что существенно различается с распределением мирных (карповых) рыб. Показано [10], что отличия и особенности вертикального распределения разновозрастных и разноразмерных особей хищных видов рыб связаны с внутренними механизмами внутривидовой конкуренции внутри каждой популяции, которые в свою очередь приводят к дифференциальной смертности среди размерных классов. Такая особенность более равномерного вертикального распределения рыб, относящихся к хищным видам, позволяет им снизить конкуренцию и агрессивное взаимодействие друг с другом. При этом в исследованиях [3] установлено, что хищные рыбы в более сложных средах проявляют меньшую агрессивность, это в свою очередь обеспечивает большую плотность рыбного населения. Водную толщу ямы, вероятно, можно отнести к таким сложным средам, поскольку здесь, помимо основного течения реки, наблюдаются постоянные восходящие потоки (с большим количеством взвешенных веществ), водовороты (суводи) и значительные перепады глубин.
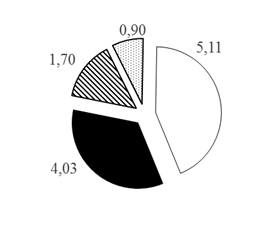
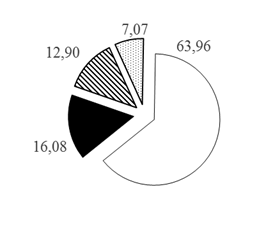
Таким образом, при рассмотрении каждого из горизонтов водной толщи по отдельности установлено, что с увеличением глубины доля карповых рыб в горизонтах глубин снижается с 80,77 до 60,26%, а доля остальных групп рыб увеличивается при движении к более глубоким горизонтам: для окуневых с 11,14 до 22,27% на глубинах >30 м, для сиговых с 5,14 до 12,90% на глубинах 20-30 м и для группы нераспознанных рыб с 2,95 до 7,86% на глубинах >30 м (рис. 3).
 А
А 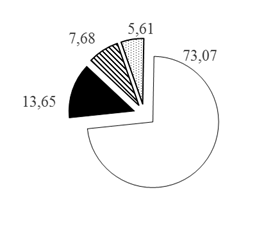 Б
Б
 В
В 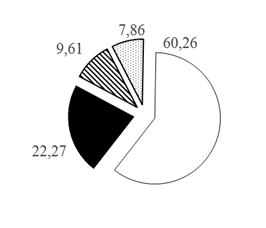 Г
Г
Рис. 3. Распределение различных таксономических групп рыб в горизонтах водной толщи Кондинской русловой ямы по горизонтам глубин (А. 0-10 м; Б. 10-20 м; В. 20-30 м; Г.>30 м), в % от общего числа зарегистрированных рыб в каждом из горизонтов: 1. Карповые 2. Окуневые 3. Сиговые 4. Нераспознанные
Особенности биотопического, а, следовательно, вертикального и горизонтального распределения рыб в водотоке являются видоспецифичными для различных видов рыб в зависимости от морфологии тела, даже у одного вида преимущественное распределение изменяется в процессе онтогенеза и размеров тела [12], физических параметров реки, например скорости течения [7]. Исследование [8] некоторых хищных видов рыб показало, что плотность жертв не имеет прямой корреляции со степенью хищничества, а наибольшее влияние на успех захвата жертвы оказывает ее визуальное обнаружение, на которое значительное влияние оказывает и мутность воды [6]; в связи с этим наибольшая часть карповых рыб распределяется в поверхностном горизонте глубин 0-10 м, где доля хищных видов – окуневых и сиговых - меньше, чем в более глубоком 10-20 м. По данным Дж. Розенфельда и Дж. Тэйлора [13], при увеличении глубины скорость течения снижается, в связи с этим растущие с возрастом рыб их потребности в энергии побуждают их перемещаться на б?льшие глубины с меньшей силой течения. По данным исследователей [14], физическими факторами, влияющими на распределение и концентрации рыб, могут быть критические скорости плавания, солнечный свет, температура, гидростатическое давление, которые в совокупности для каждого вида рыб формируют условия оптимальной биоэнергетики, вероятно, зону которой они и выбирают, тем самым распределяясь в водных экосистемах. При этом стоит учесть, что сила каждого фактора может отличаться водоема к водоему [14]. Показано [11], что вертикальное распределение рыб в водоеме обусловлено сочетанием физиологических потребностей, конкуренцией [2], наличием кормовых объектов, при этом установлено, что распределение отдельных видов по глубине схоже как для молоди, так и для взрослых особей. Изучение вертикального распределения некоторых видов сиговых рыб [9] подтверждает, что в распределение рыб значительный вклад вносит выбор зоны эффективной биоэнергетики.
Заключение. Таким образом, особенности вертикального распределения рыб в акватории русловой ямы показывают закономерности разделения мест обитания мирных и хищных видов рыб, а для хищных – равномерное распределение по горизонтам глубин, обеспечивающее снижение меж- и внутривидовой конкуренции на участках рек с русловыми ямами.
Библиографическая ссылка
Чемагин А.А. ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ РЫБ В ВОДНОЙ ТОЛЩE КОНДИНСКОЙ РУСЛОВОЙ ЯМЫ // Современные проблемы науки и образования. 2017. № 6. ;URL: https://science-education.ru/ru/article/view?id=27062 (дата обращения: 10.07.2026).