



Биогеоценоз – сложная природная система, определяемая прямыми и косвенными взаимодействиями между ее компонентами. Обладая целостностью процессов в пределах системы, биогеоценоз может рассматриваться как совокупность подсистем (фитоценотических, микоценотических, зооценотических), которые определяют совокупные, эмерджентные свойства системы. В пределах каждого биогеоценоза существует мозаичность микроместообитаний, которая внешне проявляется в неравномерности пространственного распределения организмов в лесу и их функционирования [3; 5; 6; 10]. Эти субъединицы разные авторы обозначают как тессеры (элементарные единицы лесных биогеоценозов, на уровне которых реализуются взаимосвязи растительность - почвенная биота - почва [8; 13]) или консорции [2; 11]. Более распространена концепция консортивной структуры биогеоценоза, согласно которой «каждый организм обычно входит в состав общего биоценоза не сам по себе, а в составе какого-либо консорция, состоящего из одной особи вида - эдификатора консорция и целого ряда особей - эпибионтов и эндобионтов, поселяющихся на теле эдификатора» [2, с. 7]. Л.Г. Раменский [12, с. 186-187] понимал под консорцией «сочетание разнородных организмов, тесно связанных друг с другом в их жизнедеятельности известной общностью судьбы». Чаще всего объектом исследований являются автотрофные, реже – гетеротрофные консорции, так как динамичность гетеротрофов является серьезным препятствием для мониторинга структуры консорций.
На формирование мозаичности среды биогеоценоза влияют разнообразие условий из-за специфики микро- и мезорельефа, отличия в характеристиках консорций разных детерминантов, а также специфические условия микроместообитаний, формирующихся при разложении крупных единиц древесного детрита (валежные стволы, крупные ветви, пни), древесные остатки (валежные стволы, пни и т.п.). Некоторые авторы [1; 4; 17] считают комплексы организмов, обеспечивающих разложение подобного детрита, консорциями, но в этом случае вызывает сомнение ядро консорции, которое в данном случае – не живой организм, а мортмасса, которая к тому же достаточно интенсивно исчерпывается за счет деятельности деструкторов и детритофагов разных систематических групп, причем состав этих групп постоянно меняется по мере развития процесса гниения древесины [15; 16].
На наш взгляд, с точки зрения классической трактовки консорции, древесный детрит априори не может рассматриваться в качестве детерминанта консорции; его наличие вносит свою лепту в формирование разнообразия экотопов в пределах биогеоценоза. Наша гипотеза заключается в том, что в непосредственной близости от единицы детрита формируется специфичный микроклимат, так как микогенная деструкция древесины сопровождается особым режимом температуры и влажности [7–9]. В совокупности с собственно трофическим ресурсом эти условия создают специфическую среду микроэкотопа, обеспечивающую более эффективную деятельность редуцентов [13; 14].
Материалы и методы
Для оценки специфики среды микроэкотопов, формируемых древесным детритом, в 2017 году были проведены исследования в осиннике разнотравном в пойме р. Ключ (с. Ташла Тюльганского района Оренбургской области). Производился суточный мониторинг температуры и влажности с использованием логгеров EClerk-USB-RHT с периодичностью контроля 10 минут. Логгеры закладывались в осиннике разнотравном на валежном стволе осины со следами деструкции (изменение структуры древесины, наличие белой гнили, плодовые тела трутовика настоящего – Fomes fomentarius (Fr.) Fr.) и на стволе осины без признаков деструкции (бревно осины, спиленное год назад). У каждого ствола были заложены поперечные профили с размещением логгеров с интервалом 15 см в обе стороны от каждого ствола.
Результаты и обсуждение
Анализ хода температур в биотопе (осиннике разнотравном) показал отличия в динамике температуры биогеоценоза в целом и динамике показателей деструктурируемого детрита и плодовых тел трутовиков (рис. 1).
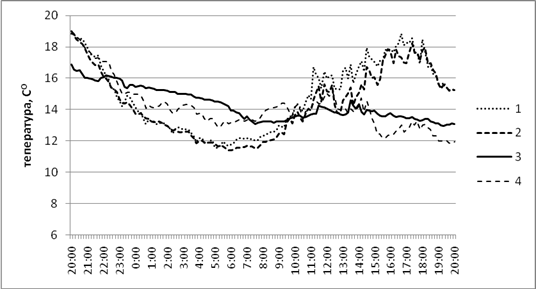
Рис. 1. Динамика температуры в осиннике разнотравном.
Положение логгеров: 1 – среда; 2 - у валежного ствола; 3 - внутри валежного ствола; 4 - внутри базидиомы трутовика
В первом случае динамика температуры преимущественно зависит от освещенности, о чем свидетельствуют максимальные показатели, приходящиеся на период времени с 11 до 18 часов. Зависимость между температурой и освещенностью в локациях, расположенных в непосредственной близости от деструктурируемого детрита и непосредственно на нем, практически отсутствует. Динамика температуры внутри валежного ствола и внутри базидиом трутовика более синхронна (коэффициент корреляции достигает 0,85). Температура плодовых тел грибов наиболее существенно снижается в самое жаркое время суток, вероятно, в результате повышения испарения.
Динамика влажности имеет сходные тенденции, хотя отличия между показателями менее выражены. Внутри разлагаемого детрита и у базидиом трутовика показатели влажности были более высокими и медленнее варьировали в течение суток (рис. 2).
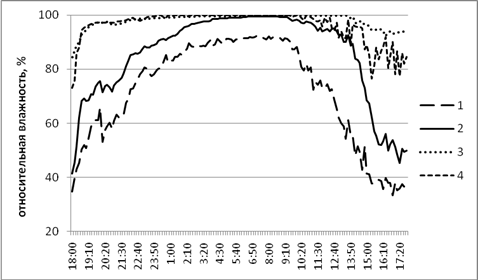
Рис. 2. Динамика влажности в осиннике разнотравном.
Положение логгеров: 1 – среда; 2 - у валежного ствола; 3 - внутри валежного ствола; 4 - внутри базидиомы трутовика
Температурно-влажностные характеристики биогеоценоза существенно отличаются в разных его частях из-за отличий в полноте древостоя, сомкнутости крон, близости опушки, определяющих освещенность, продуваемость и ряд других факторов. Как показано выше, наличие упавших стволов также вносит свой вклад в формирование мозаики микроклиматических условий биогеоценоза. Упавшие стволы деревьев, в которых не наблюдается активного процесса гниения, вероятно, формируют микроэкотопы, специфика среды которых определяется способностью объектов конденсировать влагу и снижать температуру прилегающего пространства. Было предположено, что разлагающийся детрит формирует микроэкотопы, иные по температурно-влажностным характеристикам.
Мониторинг показал, что тенденции динамики температуры и влажности вблизи разлагающегося и относительно неизмененного ствола осины существенно отличаются (рис. 3).
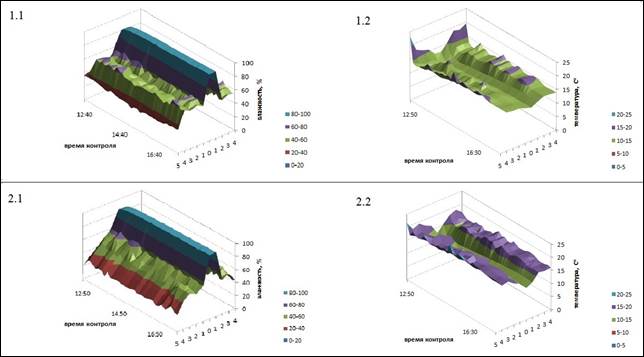
Рис. 3. Динамика показателей экотопов (1 – экотоп у ствола разлагающейся осины, 2 – у ствола осины без признаков разложения): 1.1, 2.1 – влажность; 1.2, 2.2 – температура
Поскольку оба ствола осины находились под пологом леса с сомкнутостью крон 70-80%, влияние освещенности на температуру и влажность нивелировалось. Это же касается и влияния выдувания на динамику характеристик экотона, так как стволы несущественно выделялись в рельефе и располагались на равном удалении от опушки леса.
Динамику показателей экотопа у разлагающегося ствола осины отличает общая «сглаженность» изменения показателей температуры и влажности по мере удаления от детрита; здесь же существенно ниже максимальная температура. В сравниваемом экотопе контраст в показателях по мере удаления от ствола выражен ярче; температуры на расстоянии в 20-40 см гораздо выше, чем на таком же расстоянии от гниющего ствола.
Интегральным показателем температурно-влажностных условий, отражающим испаряемость, является гидротермический коэффициент. Сравнение его динамики (рис. 4) демонстрирует тенденцию, сходную с отмеченной выше. В непосредственной близости от гниющего ствола осины более благоприятные условия для развития грибов отмечены на значительной площади и сохраняются практически неизменными на протяжении всего дня, в отличие от микроэкотопа у ствола без признаков деструкции.
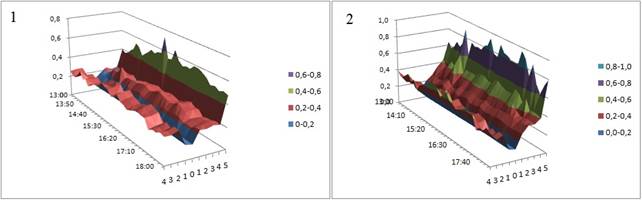
Рис. 4. Динамика гидротермического градиента в микроэкотопе разлагающегося (1) и неразлагающегося (2) детрита
Обобщая вышеприведенные факты, можно сделать вывод, что разлагающийся детрит оказывает существенное влияние на микроклиматические условия прилегающего пространства, в связи с чем формируется экотоп, специфичный с точки зрения абиотических условий. Под влиянием средообразующей деятельности разлагающегося детрита показатели температуры и влажности изменяются в течение дня более поступательно. В качестве вероятной причины такого влияния детрита на состояние среды можно рассматривать изменение режима влажности древесины из-за микогенной деструкции, в ходе которой происходит изменение структуры древесины, обводнение части тканей ствола, активное испарение влаги с поверхности плодового тела гриба, как инструмент снижения нагрева.
Формирование ксилотрофными грибами специфичных условий среды при деструкции детрита, вероятно, направлено на достижение нескольких целей. Во-первых, создающиеся условия благоприятны для роста и функционирования мицелия самого гриба; во-вторых, формирующийся микроклимат благоприятен для заселения близлежащих субстратов грибами. Другими словами, микогенная деструкция инициирует активное заселение древоразрушающими грибами соседних субстратов. Таким образом, микоценоячейки, решая свои экосистемные задачи, поддерживают существование ксиломикокомплексов в лесных экосистемах, одновременно обуславливают увеличение мозаичности условий биогеоценоза.
Библиографическая ссылка
Сафонов М.А., Остапенко А.В., Уварова А.И. СПЕЦИФИКА ЭКОТОПОВ, ФОРМИРУЕМЫХ ДРЕВЕСНЫМ ДЕТРИТОМ В ЛЕСНЫХ ЭКОСИСТЕМАХ ЮЖНОГО ПРЕДУРАЛЬЯ // Современные проблемы науки и образования. 2017. № 6. ;URL: https://science-education.ru/ru/article/view?id=27053 (дата обращения: 30.06.2026).