



Симпатическая нервная система и механизмы ее регуляции обеспечивают во многом выживание организма в различных условиях внешней среды [1]. В процессе эволюции сложились алгоритмы взаимодействия симпатической нервной системы с другими механизмами регуляции гомеостаза в организме [1, 2]. Например, стресс вызывает активацию симпатической нервной системы, и обычно при этом повышается двигательная активность. Повышение двигательной активности приводит к мышечной гиперемии, увеличению кровотока в работающих мышцах, хотя при стрессе в крови увеличивается концентрация норадреналина и адреналина, которые вызывают сокращение артерий [1-3]. Исходя из факта, что кровоток в работающих мышцах возрастает, это значит, что артерии в работающих мышцах становятся менее чувствительными к норадреналину и адреналину. Этот факт позволяет нам предположить, что сам организм вырабатывает вещества, предохраняющие артерии от излишнего сокращения, а значит, и от гипертонии. Но при развитии цивилизации значительно уменьшилась мышечная активность, а значит, и мышечная гиперемия, что привело к уменьшению и выработке сосудорасширяющих веществ. В последнее время значительно возросло количество сердечно-сосудистых заболеваний. Одной из причин этого считается гиподинамия [4], которая связана с облегчением физического труда, большую часть времени человек проводит за компьютером и телевизором. Физиологи значительное время изучали мышечную гиперемию и показали [4, 5], что при мышечной нагрузке, даже при ходьбе, резко усиливается кровоток в работающих мышцах, и мышцы даже выполняют функцию насоса, облегчая работу сердца [4]. Мышечная гиперемия способствует снижению артериального давления за счет расширения артерий, известно, что гиподинамия способствует возникновению гипертонии. Изучая мышечную гиперемию, ученые выдвинули много гипотез и теорий, доказывая различные механизмы возникновения этого феномена [6-8]. Но при анализе литературных источников мы не встретили работ по изучению и анализу количественных характеристик альфа-1-адренорецепторов артерий при мышечной гиперемии [3, 4, 9]. Следует отметить, что при гипертонической болезни у ряда больных увеличена концентрация в крови катехоламинов, поэтому они и принимают лекарства, блокирующие адренорецепторы артерий [2], что ведет к расширению артерий и снижению давления. Закономерно возникает вопрос, можно ли с помощью дозированных физических нагрузок, за счет мышечной гиперемии расширить артерии и снизить артериальное давление без лекарств. Но на этом пути возникает много вопросов, например, не изучены полностью количественные фундаментальные механизмы мышечной гиперемии. Поэтому, целью наших исследований было выявление рецепторных механизмов мышечной гиперемии, полученной при электростимуляции мышц. Исследование имеет не только фундаментальный характер, но и может помочь частично решить проблему лечения гипертонии научно подобранной для данного человека физической нагрузкой.
Методы исследования. Опыты проведены на 30 кроликах весом 2,5–3 кг под гексеналовым наркозом (внутривенно 30 мг/кг однократно). Контрольную группу составили 15 кроликов, у которых не проводили электростимуляцию мышц. Экспериментальную группу составили 15 кроликов, у которых была моделирована мышечная гиперемия методом электростимуляции мышц задней конечности. Регистрировали тонус артерий задней конечности при перфузии бедренной артерии [3] постоянным объемом крови 4,6 мл/мин (при полной перевязке анастомозов). Данные перфузионного давления с помощью датчиков преобразовывались и через аналого-цифровой преобразователь регистрировались компьютером десять раз в секунду, обрабатывались программой анализа. Те же данные регистрировались и на широкоформатном самописце. Различные дозы норадреналина (0.5 мкг/кг-1,0- 2,0- 2,5-3-5-10-15-20-30 мкг/кг) вводились в бедренную артерию перед насосом постоянной производительности, и величина повышения перфузионного давления характеризовало альфа-1-адренореактивность артерий конечности в контроле и на фоне мышечной гиперемии. По величинам доза-эффект в двойных обратных координатах Лайниувера – Берка находили количество активных альфа-1-адренорецепторов и чувствительность альфа-1-адренорецепторов [2, 3]. Количество активных адренорецепторов (Рм) было пропорционально величине повышения перфузионного давления на максимальную дозу норадреналина, дальнейшее увеличение дозы норадреналина не увеличивало давление, так как уже 100 % адренорецепторов были возбуждены. Величина чувствительности (1/К) находилась как обратная величина дозы норадреналина (К), которая численно была равна дозе норадреналина, вызывающей 50 % от максимального (Рм) эффекта повышения перфузионного давления. Этот показатель широко распространен в фармакологии и называется (ЛД50), доза, вызывающая 50 % от максимального лечебного эффекта [2, 3]. Метод анализа в двойных обратных координатах заключается в том, что максимальные величины дозы норадреналина и соответственно прессорная реакция артерий возрастают к центру (рис. 2) графика (в обычных координатах, наоборот, величины возрастают от центра). Поэтому двойные обратные координаты методом экстраполяции (рис.2) позволили получить максимально возможную величину доза-эффект (1/Рм), которая на графике расположена ближе к центру графика (в прямых координатах эти величины уходят в бесконечность). Этот метод хорошо изучен в ферментативной кинетике, биохимии, физиологии [2, 3], он применяется для анализа лиганд-рецепторных взаимодействий. Для получения рабочей гиперемии мышцы задней конечности кролика стимулировали игольчатыми электродами прямым методом частотой 5 гц при напряжении 10 вольт при длительности импульса 5 миллисекунды. После включения электростимулятора мышцы сокращались, бедренные артерии расширялись, возникала мышечная гиперемия, перфузионное давление снижалось. На фоне постоянной электростимуляции с частотой 5 гц вводили те же дозы норадреналина, что и в контроле без электростимуляции. Сравнение величин повышения перфузионного давления в артериях конечности на разные дозы норадреналина в контроле и после рабочей гиперемии позволило впервые дать количественную характеристику альфа-1-адренореактивности при мышечной гиперемии.
Для анализа данных эксперимента использованы адекватные методы статистического анализа по Стьюденту [3]. При работе с экспериментальными животными были выполнены все правила биоэтики для работы в данных условиях.
Результаты исследования и их обсуждение. Введение норадреналина в различных возрастающих дозах в бедренную артерию (рис.1) перед насосом в контрольной группе кроликов без электростимуляции всегда вызывало повышение перфузионного давления за счет возбуждения прессорных постсинаптических а1-аренорецепторов [2].
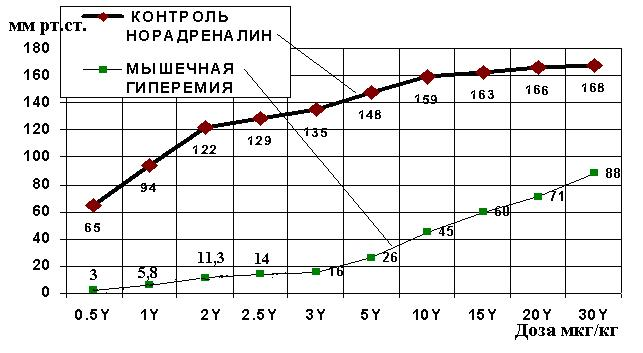
Рис. 1. Средние величины повышения перфузионного давления в бедренной артерии на различные дозы норадреналина (мкг/кг) в контроле (ромбы, верхняя линия) и после мышечной гиперемии (квадраты, нижняя линия). По оси абсцисс: дозы норадреналина в мкг/кг (Y) при внутриартериальном введении перед перфузионным насосом постоянного расхода, качающего кровь в бедренную артерию. По оси ординат: повышение перфузионного давления в мм рт. ст. на норадреналин после мышечной гиперемии. Разница всех величин реакций контрольной группы и после мышечной гиперемии имеют статистически значимые различия (P<0.05)
Увеличение дозы норадреналина всегда увеличивало прессорную реакцию артерий и усиливало повышение перфузионного давления в задней конечности. Проведенные исследования показали, что у контрольных животных было намного большее повышение перфузионного давления на норадреналин, чем введение норадреналина на фоне электростимуляции мышц при (рис.1) мышечной гиперемии.
На фоне сокращения мышц при мышечной гиперемии введение норадреналина вызывало гораздо меньшее сокращение артерий конечности на все исследуемые дозы. Прессорное действие норадреналина (рис.1) на артерии конечности кролика при мышечной гиперемии наиболее было выражено при низких дозах норадреналина, при увеличении доз норадреналина это различие уменьшалось.
При дозе вводимого норадреналина (рис.1) 0,5 мкг/кг в перфузируемую бедренную артерию в контроле давление увеличилось на 65 мм рт. ст. от исходного уровня, а при мышечной гиперемии только на 3 мм рт. ст. Значит, во время мышечной гиперемии прессорное действие норадреналина уменьшилось в 22 раза. При дозе норадреналина 1 мкг/кг в контроле давление возросло на 94 мм рт. ст., а во время мышечной гиперемии увеличилось только на 5,8 мм рт ст., то есть было в 16 раз меньше. При дозе норадреналина 2 мкг/кг в контроле давление возросло на 122 мм рт. ст., а во время мышечной гиперемии увеличилось только на 11,3 мм рт ст., то есть было в 10,8 раза меньше. При дозе норадреналина 2,5 мкг/кг в контроле давление возросло на 129 мм рт ст., а во время мышечной гиперемии увеличилось только на 14 мм рт. ст., то есть было в 9,2 раза меньше. При дозе норадреналина 5 мкг/кг в контроле давление возросло на 148 мм рт. ст., а во время мышечной гиперемии увеличилось только на 26 мм рт. ст., то есть было в 5,7 раза меньше. При дозе норадреналина 10 мкг/кг в контроле давление возросло на 159 мм рт ст., а во время мышечной гиперемии увеличилось только на 45 мм рт. ст., то есть было в 3,5 раза меньше. При дозе норадреналина 30 мкг/кг в контроле давление возросло на 168 мм рт. ст., а во время мышечной гиперемии увеличилось только на 88 мм рт. ст., то есть было в 1,9 раза меньше.
Для установления механизмов этого различия действия норадреналина на артерии в контроле и после мышечной гиперемии (рис. 2) был проведен кинетический анализ действия норадреналина в двойных обратных координатах Лайниувера – Берка, который позволил определить максимально возможную прессорную реакцию (Рм) артерий на норадреналин, которая пропорциональна количеству активных адренорецепторов артерий. Получено (рис.2), что количество активных адренорецепторов к норадреналину до и после мышечной гиперемии не изменилось, так как максимальная прессорная реакция на норадреналин была одинакова до и после гиперемии (Рм=172 мм рт. ст.). Этим же методом, Лайниувера – Берка, была определена чувствительность реакции медиатор-рецептор (1/К) до и после гиперемии.
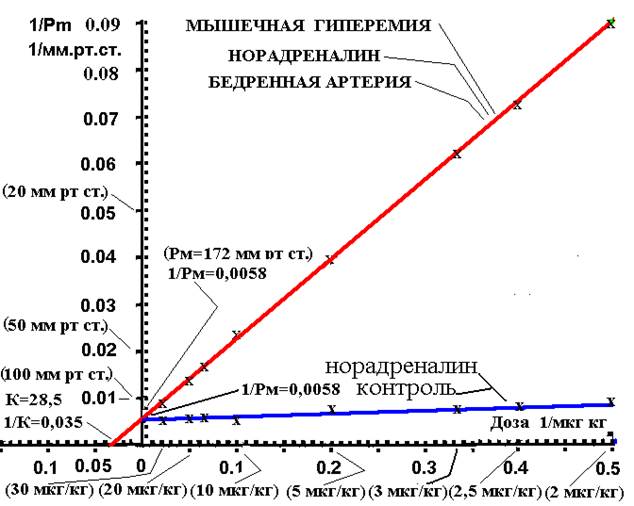
Рис. 2. Изменение реактивности а1-адренорецепторов артерий к различным дозам норадреналина в двойных обратных координатах Лайниувера – Берка в контрольной группе и на фоне мышечной гиперемии. По оси абсцисс: обратная величина дозы норадреналина (1/Доза в 1/(мкг/кг) при внутриартериальном введении. По оси ординат: обратная величина (1/Рм) повышения перфузионного давления в (1/мм рт ст.) на норадреналин. Красная линия – опыты на фоне мышечной гиперемии. Синяя линия – опыты в контрольной группе. Разница всех величин реакций контрольной группы и после мышечной гиперемии достоверны (P<0.05). Данные доз норадреналина 0,5 и 1 мкг/кг и прессорной реакции лежат на продолжении прямых линий доза-эффект, на графике (рис. 2) они не представлены, так как ось абсцисс тогда была бы в 3 раза длиннее
Построение графиков доза-эффект в двойных обратных координатах Лайниувера – Берка позволило методом экстраполяции (рис. 2) определить максимально возможную Рм=172 мм рт ст. прессорную реакцию (1/Рм-обратная величина) артерий на самую большую дозу норадреналина. Значит, что в контрольной группе и после мышечной гиперемии перфузионное давление в артериях конечности может повышаться максимально на 172 мм рт. ст. на максимально большие дозы норадреналина. Это доказывает, что количество активных альфа-1-адренорецепторов до и на фоне мышечной гиперемии не изменилось.
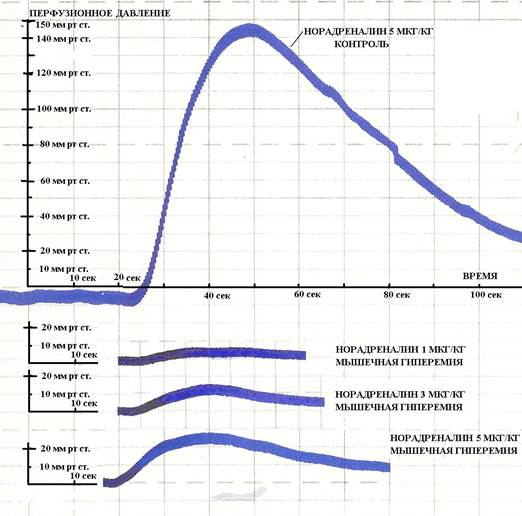
Рис. 3. Повышение перфузионного давления в бедренной артерии после введения различных доз норадреналина в контрольной группе животных и на фоне мышечной гиперемии. По оси абсцисс время реакции в секундах. По оси ординат величина прессорной реакции перфузионного давления в мм рт ст. бедренной артерии после введения норадреналина. (Фото с ленты самописца)
Изучение физиологических механизмов уменьшения реактивности адренорецепторов артерий к норадреналину при рабочей гиперемии показало (рис.1, 2, 3), что только за счет уменьшения величины чувствительности (в контроле 1/К=1,2; после мышечной гиперемии 1/К=0,035) альфа-1-адренорецепторов к норадреналину уменьшается его прессорное действие на артерии. Анализ максимально возможной прессорной реакции артерий на норадреналин, которая характеризует количество активных адренорецепторов артерий, показал, что количество активных адренорецепторов после рабочей гиперемии не изменилось. Такое уменьшение чувствительности а1-адренорецепторов к норадреналину при мышечной гиперемии при одинаковом их количестве с контролем и обеспечивает механизм, при котором при низких дозах норадреналина величина прессорной реакции при мышечной гиперемии снижается гораздо больше, чем при больших дозах норадреналина.
Заключение. Изучение эффекта мышечной гиперемии у животных и человека в основном было направлено исследователями на величину увеличения кровотока при сокращении мышц и изучении биохимического состава крови [1, 6, 7]. Но при анализе публикаций во всех доступных базах данных мы не встретили ни в одной работе данных о влиянии мышечной гиперемии на альфа-адренореактивность артерий. Поэтому полученные нами результаты частичного блокирования альфа-адренорецепторов при мышечной гиперемии вносят вклад в развитие фундаментальных знаний в области физиологии человека и животных и являются основанием для проведения исследований в этой области, конечным результатом которых должны стать клинические рекомендации по использованию дозированной физической нагрузки у пациентов с артериальной гипертензией. Наши опыты показали, что на все дозы норадреналина прессорная реакция бедренной артерии кролика намного меньше при мышечной гиперемии (рис.1). В результате проведенного исследования установлено, что мышечная гиперемия, вызванная электрической стимуляцией мышц, вызывает расширение артерий за счет значительного уменьшения чувствительности альфа-1-адренорецепторов артерий (рис. 2).
Установлено, что при использовании низких доз норадреналина разница в реактивности артерий к норадреналину в контроле и на фоне мышечной гиперемии более выражена, чем при использовании в эксперименте высоких доз. Установлено, что такое различие реактивности артерий к различным дозам норадреналина в контроле и на фоне мышечной гиперемии обусловлено тем, что количество альфа-1-адренорецепторов артерий не изменилось, но значительно снизилась их чувствительность на фоне мышечной гиперемии.
Библиографическая ссылка
Ананьев В.Н., Демидов В.А., Ананьев Г.В. РЕАКТИВНОСТЬ К НОРАДРЕНАЛИНУ АРТЕРИЙ ЗАДНЕЙ КОНЕЧНОСТИ КРОЛИКА ПРИ РАБОТАЮЩИХ МЫШЦАХ // Современные проблемы науки и образования. 2017. № 4. ;URL: https://science-education.ru/ru/article/view?id=26601 (дата обращения: 02.07.2026).