



Мясная продуктивность крупного рогатого скота имеет важное значение в отрасли животноводства, особенно в получении качественной продукции.
Развитие молекулярной биологии предопределило развитие такой отрасли, как маркерная селекция. Этому способствовал прогресс в понимании представлений о путях и методах исследования различных процессов в организме животного, которые обусловлены не только действием генов, но и факторами внешней среды [1-3]. Так, правильное выращивание и откорм молодняка определяют уровень мясной продуктивности в равной степени с активностью генов [4]. Особое значение данные процессы играют в селекции, которая направлена на получение животных с хозяйственно ценными признаками.
Использование ДНК-маркеров позволяет оценить гены, играющие значительную роль в формировании и регуляции ряда физиологических процессов. Поскольку ген обладает различными аллельными вариантами (полиморфизмы), связанными с вариативностью уровня продуктивности, то «считывание» этих вариантов позволит выявить желательные сочетания аллелей и позволят провести селекцию животных по генотипам [4; 5].
В раннее проведенных исследованиях по оценке влияния полиморфизма гена CAPN1, который обуславливает модификацию мышечной структуры, была выявлена не только зависимость структурно-механических свойств мяса от наличия или отсутствия желательного аллеля, но и изменение аминокислотного состава мяса на примере валина. Этот факт позволил констатировать, что наличие этого гена в желательной аллельной форме способствует изменению в белковом синтезе [6]. Полученные данные позволили предположить, что животные, имеющие данную мутацию, будут характеризоваться более ускоренными темпами роста при стандартном кормлении. Это обусловлено тем, что данная аминокислота играет ключевую роль в метаболизме мышц и восстановлении поврежденных тканей [7; 8].
Таким образом, изучение биологических и генетических закономерностей, влияющих на процессы роста и развития животных крупного рогатого скота, а также возможность применения полученных знаний в практике современного животноводства и селекционного процесса позволит получить животных с новыми качественными хозяйственно полезными признаками [9].
Исходя из вышесказанного, целью нашего исследования была оценка влияния наличия полиморфизма гена CAPN1 на рост и развитие животных при условиях стандартного кормления.
Материалы и методы
Хозяйственный эксперимент был проведен в условиях ООО «Агробизнес» Республики Калмыкия, лабораторные исследования – на базе испытательного центра Всероссийского научно-исследовательского института мясного скотоводства.
Для прижизненной оценки взаимосвязи влияния различных аллельных вариантов гена CAPN1 на рост и развитие была использована кровь бычков калмыцкой породы крупного рогатого скота (КРС) (n=70). Исследование проводилось с использованием ПЦР в реальном времени.
Выделение ДНК из крови проводили с использованием комплекта реагентов для выделения геномной ДНК из цельной крови «ДНК-Экстран-1» («Синтол», Россия).
Для амплификации были использованы праймеры, последовательность которых соответствовала фрагменту гена CAPN1 (5’-AGCAGCCCACCATCAGAGAAA – 3’; 5’- TCAGCTGGTTCGGCAGAT – 3’).
Объем реакционной смеси составил 25 мкл и содержал 60 мМ трис-HCl (рН 8,5), 1,5 мМ MgCl2, 25 мМ KCl, 10 мМ меркаптоэталол; 0,1 мМ тритон Х-100; 0,2 мМ дНТФ, 1 ед. Taq ДНК полимеразы, по 0,2 мкМ каждого из праймеров. Амплификацию гена CAPN1 проводили на программируемом амплификаторе АНК-32 («Синтол», Россия) по следующему режиму:
1. 95 ºС – 120 сек х 1;
2. 64 ºС – 40 сек х 40;
3. 95 ºС - 20 сек х 40.
На основании результатов лабораторных испытаний по анализу наличия полиморфизма по гену CAPN1 из общего числа животных было сформировано 3 группы животных (n=15): I – без мутационной аллели (GG); II – гетерозиготное ее проявление (GC); III – с наличием полиморфизма в гене CAPN1 (CC).
На следующем этапе была проведена оценка особенностей роста и развития подопытных животных с целью оценки наличия взаимосвязи между исследуемыми параметрами.
Кормление животных осуществлялось в соответствии с нормами, предусматривающими один и тот же уровень кормления, рассчитанный на получение 900-1000 г среднесуточного прироста (Калашников А.П. и др., 2003). Уровень кормления во всех сравниваемых группах был одинаковым и вполне соответствовал потребностям растущего молодняка.
Контроль роста подопытных животных проводился путем ежемесячного индивидуального взвешивания (утром до кормления). На основании этих данных были рассчитаны среднесуточный прирост массы тела, относительная скорость роста в отдельные возрастные периоды.
Относительная скорость роста подопытных животных вычислялась по формуле С. Броди:
![]() ,
,
где К – относительная скорость роста;
W1 – начальная живая масса, кг;
W2 – конечная живая масса, кг.
Учет проводился до 14 месяцев.
Статистическая обработка проводилась с применением общепринятых методик при помощи приложения Statistica 10.0, включая определение средней арифметической величины (М), стандартной ошибки средней (m). Достоверными считали различия при р≤0,05. Оценку статистической значимости различий между группами проводили с помощью t-критерия Стьюдента (О.Ю. Реброва, 2002).
Результаты исследования
В результате проведенных исследований выявлено, что наличие полиморфизма по гену CAPN1 влияет на ростовые характеристики животных.
Анализ данных показал, что различия в массе наблюдаются, начиная с 3 месяцев между животными III и I групп, при рождении животные всех исследуемых групп характеризовались сходной живой массой. Так, в III группе живая масса в этом возрасте была на 2,2% и 1,4% больше, чем в I и II группах соответственно. Живая масса во II группе, в сравнении с I, была больше на 1,8%.
С увеличением возраста животного разница по массе продолжала увеличиваться. В 8 месяцев животные III группы имели в среднем живую массу 227,7 кг, в то время как животные I и II групп отставали по данному показателю на 8,4% (P <0,05) и 4,0% соответственно. Разница между I и II группами была незначительной.
В годовалом возрасте разница составила 6,3% и 3,2% соответственно. На момент окончания эксперимента разница между I и III группами была 6,2%, а между III и II – 3,5% (рис. 1).
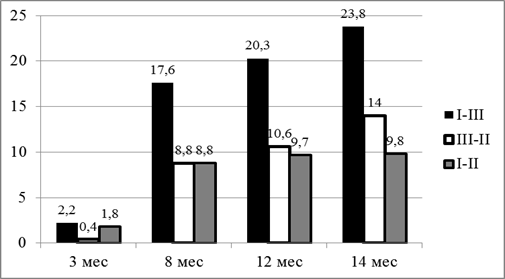
Рис. 1. Динамика разницы в живой массе между сравниваемыми группами, кг
Анализ динамики среднесуточных приростов показал, что максимальный прирост наблюдался период c 13-го по 14-й месяц, причем распределение между группами было не одинаковым. Так, в I группе этот показатель составил 1042 г, во II – 1047 г, а в III – 1082 г. Причем следует отметить, что разница между I и II группами была незначительной, а животные III группы по данному показателю превышали своих сверстников из I группы на 13% (Р>0,99), из II – 12,6% (таблица 1).
Таблица 1
Динамика среднесуточных приростов бычков разных генотипов, г
|
Возрастной период, мес. |
Группа |
||
|
I |
II |
III |
|
|
0-8 |
734±3,78 |
773±6,26 |
809±6,51 |
|
9-12 |
939±3,16 |
945±7,09 |
960±2,70 |
|
13-14 |
1042±6,01 |
1047±11,4 |
1082±3,81 |
Расчет абсолютного прироста животных показал, что животные III группы характеризовались более высокими результатами в сравнении с I и II. Так, в период с 7-го по 8-й месяц этот показатель в группе с генотипом GG был равен 20,2 кг, разница составила между группами GG и СС – 1,9%, и 1,7% - между животными с генотипами CC и GC. С увеличением времени разница в данных эколого-генетических группах также увеличивалась.
За весь учетный период разница в абсолютном приросте составляла в группах GG-CC, CC-CG и GG-GC 13,42, 9,67 и 3,45 кг соответственно Максимальные значения прироста были достигнуты в периоды 11-12 месяцев и 13-14 месяцев (рис. 2).
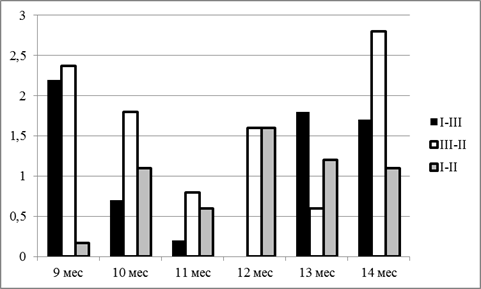
Рис. 2. Динамика разницы в абсолютном приросте животных сравниваемых групп, кг
За весь учетный период разницы в абсолютном приросте составляли в парах групп I-III, III-II и I-II – 5,6, 5,2 и 0,4 кг соответственно. По итогам исследований нами была проведена оценка относительной скорости роста бычков (таблица 2).
Таблица 2
Динамика относительной скорости роста подопытных бычков, %
|
Возрастной период, мес. |
Группа |
||
|
I |
II |
III |
|
|
0-8 |
144 |
147 |
149 |
|
9-12 |
42 |
41 |
40 |
|
13-14 |
18 |
17 |
18 |
Различия между группами по величине относительной скорости роста имели место на протяжении всего эксперимента, причем наиболее значительными были в период до 8 месяцев. Закономерно, что величина данного показателя в III группе оказалась наибольшей и превышала значение этого показателя на 5% в I и на 2% - во II группе. Между тем в последующие периоды: 9-12 и 13-14 месяцев, I группа не уступала аналогам из II и III групп. Это развитие событий связано с меньшей живой массой животных I группы в начале оцениваемых периодов. В конечном итоге при оценке динамики роста предпочтение отдается абсолютным, а не относительным величинам.
Анализируя гомо- и гетерозиготное проявление гена, следует отметить, что явная разница в показателях была отмечена непосредственно в группе животных-носителей генотипа CC. Разница между GG и GC была незначительной. Данный факт можно объяснить тем, что именно в состоянии CC данный полиморфизм оказывает видимое влияние на ростовые характеристики.
В научной литературе ген CAPN1 определяется как ген, кодирующий комплекс протеолитических ферментов, которые определяются как фактор, инициирующий декомпозицию мышечных тканей и волокон. В ходе исследований было определено существование двух основных типов кальпаинов, каждый из которых проявляет свою активность при различных концентрациях Ca2+ . Так, кальпаин A, или μ-кальпаин, активен при концентрации Ca2+ 50-100 мкМ, а кальпаин В, или М-кальпаин, активен при концентрации Ca2+ 1-2 мкМ [10].
В функциональном отношении было выявлено, что именно кальпаиновая протеаза играет ключевую роль в улучшении качества мяса за счет действия белков титина и тубулина. Результаты молекулярно-генетических исследований, в частности секвенирования II регуляторной субъединицы гена CAPN1, показали важность полиморфизма этого участка генома. В практическом отношении проведен ряд исследований, доказывающих, что наличие данного полиморфизма у сельскохозяйственных животных способствует формированию качественно полезного признака, а именно нежность мяса [11].
Параллельно с этим проводятся дополнительные исследования по выявлению зависимости ряда других признаков от наличия полиморфизма гена CAPN1, в частности качества мяса после убоя или особенности роста животного. В работах Chung et al. было показано, что точечные мутации в участке гена кальпаина III овцы влияют на формирование жировой прослойкой вокруг бедер и внутренних органов [12]. В исследованиях, проведенных на КРС, были получены результаты, доказывающие взаимосвязь с такими свойствами, как рост, динамика массы и т.д. Полученные нами данные по анализу динамики роста подопытных бычков наглядно демонстрируют, что между носительством мутации CAPN1 и интенсивностью роста есть тесная связь, а гомозиготное состояние гена С находится в определенной связи с повышенной энергией роста.
Исходя из этого, продолжение исследований в данной области представляется достаточно перспективным и требует дальнейшего изучения.
Работа выполнена при финансовой поддержке Министерства образования (Грант Президента-МК-3631.2017.11)
Библиографическая ссылка
Косян Д.Б., Русакова Е.А., Кван О.В. ОЦЕНКА ВЗАИМОСВЯЗИ ПОЛИМОРФИЗМА ГЕНА CAPN1 С РОСТОВЫМИ ХАРАКТЕРИСТИКАМИ КРУПНОГО РОГАТОГО СКОТА // Современные проблемы науки и образования. 2017. № 2. ;URL: https://science-education.ru/ru/article/view?id=26320 (дата обращения: 04.07.2026).