



Сравнительно-этологическое исследование стереотипов, повторяющихся у разных видов животных, позволяет развить представление об эволюционном формировании и путях адаптации поведения [1]. Хорошим модельным примером служит стереотип охотничьего поведения у грызунов. Среди них есть как специализированные охотники – представители рода кузнечиковых хомячков Onychomys, обладающие морфологическими и физиологическими адаптациями [8], так и эврифаги, проявляющие охотничье поведение факультативно, например оленьи хомячки (Peromyscus maniculatus) [6]. Недавно обнаруженное охотничье поведение у полевых мышей (Apodemus agrarius) [7] и рыжих полевок (Myodes glareolus) [5], не имеющих морфологических или физиологических адаптаций к охоте, позволяет предположить наличие у них специфических поведенческих приспособлений. Возникает вопрос о наличии у грызунов скрытых стереотипов поведения, использование которых расширяет адаптивный потенциал вида, за счет возможности переключения на животную пищу.
Цель данной работы – исследовать охотничье поведение у грызунов с различной пищевой специализацией в сравнении с представителем насекомоядных (обыкновенной бурозубкой).
Материалы и методы
Исследования проводились в 2012–2015 гг., в лаборатории на всеядных грызунах – серой крысе (Rattus norvegicus) (n=81), хомячке Кэмпбелла (Phodopus campbelli) (n=19), зерноядной полевой мыши (A. agrarius) (n=26), зеленоядной узкочерепной полевке (Lasiopodomys gregalis) (n=43) и обыкновенной бурозубке (Sorex araneus) (n=11). Все особи серой крысы и хомячков Кэмпбелла, а также 9 полевых мышей родились в лаборатории и до тестирования не имели опыта охоты, остальные животные были отловлены в естественных условиях (Новосибирская область). Зверьки содержались в индивидуальных клетках и имели постоянный доступ к воде и пище.
Для наблюдений животных по одному помещали в прозрачную арену (45×45×50 см для крыс, 30×30×35 см для остальных). Спустя 5 мин в арену помещали добычу – мраморного таракана (Nauphoeta cinerea) (средняя длина тела 27,93±0,22 мм). Если животное охотилось, то на арену помещали последовательно еще двух тараканов, если не проявляло интереса к добыче, то наблюдения прекращали спустя 10 минут. Серые крысы тестировались один раз. Поскольку особей остальных видов было меньше, чем крыс, то для получения сопоставимого количества стереотипов мы увеличили для них количество тестов: полевых мышей и обыкновенных бурозубок тестировали два раза, узкочерепных полевок – три, а хомячков Кэмпбелла семь раз. Действия животных фиксировались с помощью видеокамер: Sony Handycam DCR-SR68 (крысы и мыши), Sony HDR-AS200V со скоростью записи 60 кадров в секунду (полевки, хомячки и бурозубки). Анализировались только случаи успешной охоты, закончившиеся поимкой и поеданием добычи.
Для описания поведения в качестве элементарной единицы нами выделяются элементарные двигательные акты и позы («элементы поведения»). Поведенческой последовательностью мы называем произвольный набор последовательно совершаемых элементов поведения. Внутри поведенческих последовательностей мы выделяем поведенческие стереотипы, состоящие из устойчиво повторяющихся «цепочек» элементов поведения (подробно см.: [2]). Всего было выделено 19 элементов поведения, которые мы разделили на 3 типа. «Ключевые», без которых совершение стереотипа невозможно: преследование добычи бегом (Q) или спокойным шагом (S), укус (W), захват добычи лапами (E) (только у грызунов). «Дополнительные» элементы («приготовления» к охоте и поеданию добычи) присутствовали не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G), перехват (R) (только у грызунов), откусывание конечностей добычи (H) и придерживание добычи одной (N) или двумя лапами (M) – только у бурозубки. «Шумовые элементы» (не влияющие на совершение стереотипа): замирание (C), поворот корпуса на 90° (V), разворот корпуса 180° (B), поворот головы (F), вертикальная стойка (I), стойка с опорой на арену (Y), движения назад (U), прыжок (J) и чистка (X).
Обработка видеозаписи проводилась с 25-кратным замедлением в программе The Observer XT 10 (Noldus Information Technology). Используя полученный «алфавит» из 16 элементов, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Полученные поведенческие последовательности охотничьих стереотипов перемещались в отдельные для каждого вида «суммарные» текстовые файлы (в формате .txt).
Сравнения долей охотившихся и не охотившихся особей у разных видов, а также успешных и неуспешных атак у разных особей проводились с помощью точного теста Фишера. Анализ количества элементов в стереотипах (длина стереотипа) проведен с помощью H-критерия Краскела-Уоллиса; при сравнении представлены медиана, первый и третий квартили (Me; Q1 – Q3). Для построения схемы стереотипа, используя «суммарные» текстовые файлы, рассчитывали матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка) [3].
Результаты
При первом предъявлении подвижной добычи охотничье поведение продемонстрировали 67,9% (55 из 81) особей серой крысы, 65,4% (17 из 26) полевых мышей, 18,5% (9 из 46) узкочерепных полевок, 36,8% (7 из 19) хомячков Кэмпбелла и 100% обыкновенных бурозубок (n=11). После проведения повторных тестов количество особей, проявивших охотничье поведение, возросло до 80,8% (21 из 26) у полевых мышей, 39,1% (18 из 46) у узкочерепных полевок и 63,2% (12 из 19) у хомячков Кэмпбелла. По результатам всех тестов узкочерепные полевки охотились реже, чем другие виды (p < 0,01 для всех случаев).
Всего было зафиксировано 125 успешных и 77 неуспешных случаев охоты у крыс, 83 и 39 у мышей, 34 и 92 у полевок, 43 и 42 у хомячков, 61 и 34 у бурозубок соответственно. Успешность атак у узкочерепной полевки оказалась ниже, чем у всех остальных (p < 0,01 для всех случаев).
По длине охотничьи стереотипы у серых крыс (16; 9 – 28), полевок (9; 5 – 43) и бурозубок (22; 13 – 34) достоверно не различались между собой (H = 5,5, NS), но были короче, чем у мышей (31; 19 – 56) (H = 35,1, p < 0,01). Длины стереотипов хомячков (22; 13 – 34) и бурозубок (H = 3,6, NS), хомячков и мышей (H = 3,6, NS) не различались, но стереотипы хомячков были длиннее, чем у крыс (H = 12,4, p < 0,01) и полевок (H = 7,4, p < 0,01).
Скорость охоты (отношение между длиной стереотипа и его продолжительностью) у разных видов отличалась. Обыкновенные бурозубки демонстрировали (2,9; 1,8 – 4,2) элементов поведения в секунду, что достоверно больше, чем у крыс (1,2; 0,9 – 1,5), мышей (2,1; 1,5 – 2,8), полевок (1,6; 1,2 – 2,2) (H = 103,9, p < 0,01) и хомячков (1,4; 1 – 1,9) (H = 35,2, p < 0,01). Скорость охоты у полевых мышей была выше, чем у узкочерепных полевок (H = 7,3, p < 0,01) и хомячков Кэмпбелла (H = 5,4, p < 0,05). У серых крыс этот показатель оказался наименьшим среди исследованных видов: мышей (H = 57, p < 0,01), полевок (H = 8,9, p < 0,01) и хомячков (H = 2,3, p < 0,05).
Схемы стереотипов охотничьего поведения представлены на рис. 1, где показаны все устойчивые связи между элементами (вероятность перехода от одного элемента к другому p ≥ 0,2) и некоторые неустойчивые (p < 0,2), но важные для совершения стереотипа связи.
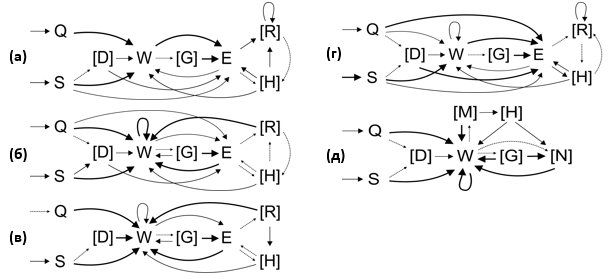
Рис. 1. Схемы охотничьих стереотипов серой крысы (а), полевой мыши (б), узкочерепной полевки (в), хомячка Кэмпбелла (г) и обыкновенной бурозубки (д)
Обозначения: тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (p < 0,2). Простой линией обозначены устойчивые связи (0,2 ≤ p < 0,5). Жирной линией обозначены высоко устойчивые связи между элементами (p ≥ 0,5). В квадратных скобках указаны дополнительные элементы.
Результаты количественного анализа представлены на рис. 2. Стереотипы охотничьего поведения полевой мыши содержали больше ключевых элементов поведения «укус» (20; 10 – 30), чем у других видов: крыс (5; 2 – 8) (H = 87,5, p < 0,01), полевок (6; 2 – 20,5) (H = 87,5, p < 0,01), хомячков (8; 4,5 – 8) (H = 17,7, p < 0,01) и бурозубок (15; 8 – 22) (H = 14,2, p < 0,05). Бурозубки кусали добычу чаще, чем крысы (H = 54,7, p < 0,01), полевки (H = 7,8, p < 0,01) и хомячки (H = 7,8, p < 0,01). У полевок и крыс, хомячков и полевок этот показатель достоверно не различался (H = 0,4, NS; H = 1, NS). Хомячки чаще хватали добычу передними лапами, чем другие исследованные грызуны: крысы (H = 14, p < 0,01), мыши (H = 5, p < 0,05), полевки (H = 42,7, p < 0,01). Полевки хватали добычу лапами реже, чем крысы (H = 4,1, p < 0,05) и мыши (H = 9,9, p < 0,01), при этом мыши захватывали добычу лапами чаще, чем крысы (H = 4,9, p < 0,05). Наибольшее количество дополнительных элементов поведения «перехват добычи» наблюдалось в стереотипах крыс (3; 1 – 4) (H = 78,5, p < 0,01), а наименьшее у полевок. Количество актов откусывания конечностей пойманной добычи в стереотипах хомячков Кэмпбелла (3; 2 – 6) выше, чем у других видов: крыс (0; 0 – 1) (H = 65,2, p < 0,01), (1; 0 – 2) мышей (H = 33,8, p < 0,01), полевок (3; 2 – 6) (H = 14,4, p < 0,01) и бурозубок (1; 0 – 3) (H = 21,2, p < 0,01).
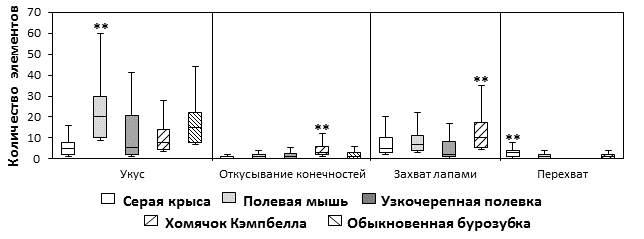
Рис. 2. Количество различных элементов поведения в одном охотничьем стереотипе. Обозначены некоторые наиболее важные различия (H-тест, ** p < 0,01)
Для дальнейшего анализа укусы были разделены на 3 группы: одиночные, двойные и множественные (3 и более укусов подряд). Отличительная особенность стереотипов крыс в том, что в них присутствовали практически только одиночные укусы, в то время как в стереотипах мышей, полевок, хомячков и бурозубок присутствовали двойные и множественные укусы (рис. 3). Количество одиночных укусов в охотничьих стереотипах бурозубок (1; 0 – 3) достоверно меньше, чем в стереотипах остальных видов: крыс (4; 2 – 7) (H = 38,6, p < 0,01), мышей (4; 3 – 7) (H = 36,4, p < 0,01), полевок (2; 1 – 7) (H = 5,7, p < 0,05) и хомячков (6; 2,5 – 10) (H = 32,2, p < 0,01). В стереотипах крыс и мышей (H = 0,8, NS), мышей и хомячков (H = 2,6, NS) количество одиночных укусов не различалось. При этом хомячки совершали одиночные укусы чаще, чем крысы (H = 4,4, p < 0,05), а те, в свою очередь, совершали одиночные укусы чаще, чем полевки (H = 4,2, p < 0,05). Среди грызунов узкочерепные полевки реже всего наносили добыче одиночные укусы. В стереотипах полевок количество одиночных укусов меньше, чем в стереотипах мышей (H = 4,9, p < 0,05) и хомячков (H = 6,5, p < 0,05). У крыс двойные и множественные укусы практически отсутствовали. Количество двойных укусов в стереотипах мышей (1; 0 – 3), полевок (1; 0 – 3), хомячков (1; 0 – 1,5) и бурозубок (1; 0 – 1) достоверно не различалось (H = 7,4, NS). В стереотипах мышей (3; 1 – 4) и бурозубок (2; 1 – 3) количество множественных укусов достоверно не различалось между собой, но было выше, чем у полевок (0; 0 – 1,8) (H = 16,6, p < 0,01; H = 12,5, p < 0,01) и хомячков (0; 0 – 0,5) (H = 43,3, p < 0,01; H = 37,6, p < 0,01) соответственно. Полевки чаще, чем хомячки, наносили добыче множественные укусы (H = 4,1, p < 0,05).
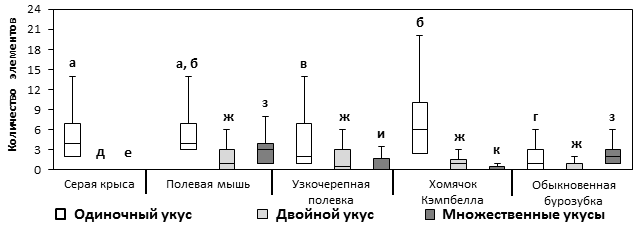
Рис. 3. Количество одиночных, двойных и множественных укусов в одном охотничьем стереотипе. Указаны медианы, квартили 25-75%, отрезки – размах. Данные, отмеченные буквами (в группах: а, б, в, г; д, ж; е, з, и, к), достоверно различаются
Обсуждение и заключение
Внимание исследователей к факультативному охотничьему поведению зерноядных грызунов было привлечено совсем недавно, а зеленоядные в этом плане практически не исследовались, за исключением нескольких видов эврифагов, в различной степени тяготеющих к питанию растительной пищей [9]. Эффективное охотничье поведение у зерноядных, с детальным описанием стереотипа охоты, было впервые выявлено у полевой мыши [7]. На примере другого, столь же широко распространенного и многочисленного вида – рыжей полевки (M. glareolus), было показано, что отбор успешных охотников в лаборатории приводит к быстрому, в течение нескольких поколений, повышению частоты проявления охотничьего поведения, что служит примером «быстрой эволюции» [5]. Характер поведения в этой работе описан не был. Мы впервые провели детальное сравнение охотничьего поведения у представителей зерноядных (полевая мышь), зеленоядных (узкочерепная полевка) и всеядных (серая крыса, хомячок Кэмпбелла) грызунов с истинным хищником, облигатно проявляющим охотничье поведение – обыкновенной бурозубкой.
У зеленоядной узкочерепной полевки – самая меньшая среди исследованных видов доля охотящихся особей и наиболее низкая успешность охоты. Примечательно, однако, что у тех особей, которые охотились, схема охотничьего стереотипа практически не отличалась от схемы стереотипа у полевых мышей. Это позволяет предположить у зеленоядных полевок наличие «скрытого» стереотипа охотничьего поведения, типичного для охотящихся грызунов.
У исследованных грызунов схемы охотничьего стереотипа по порядку совершения ключевых элементов оказались сходными, как и сам характер охотничьего поведения. Существенное различие между охотничьими атаками грызунов и насекомоядных состоит в том, что грызуны после захвата добычи зубами («укуса») осуществляют захват лапами, а бурозубки используют только зубы. Примечательно, что бурозубки атакуют добычу любых размеров только быстрыми укусами, и это считается более примитивной чертой по сравнению с захватом лапами – относительно недавним достижением в филогенетическом плане [4]. Одним из показателей большей специализации кузнечикового хомячка, как грызуна – хищника, является начало атаки с захвата добычи лапами [6]. В нашем исследовании полевые мыши и узкочерепные полевки, в отличие от кузнечиковых хомячков, начинали атаку с захвата насекомого зубами (укуса), а затем хватали его лапами. У хомячков Кэмпбелла стереотип мог начинаться как с укуса, так и с захвата добычи лапами. Таким образом, их охотничий стереотип занимает промежуточное место между другими исследованными грызунами, с одной стороны, и специализированными хищными кузнечиковыми хомячками - с другой.
Результаты сравнительного анализа длин стереотипов и скорости охоты характеризуют бурозубку как эффективного специализированного хищника. Серая крыса также естественно выступает как хищник-генералист, не проявляющий специализированного охотничьего поведения. Крысы используют укус для удержания добычи и последующего захвата лапами, они активно манипулируют удерживаемой добычей, но не умерщвляют ее, а поедают живьем, поэтому добыча часто вырывается, и зверек вынужден возобновить преследование. Хомячки Кэмпбелла эффективно обездвиживают насекомое, быстро откусывая у него все конечности. Примечательно, что как полевые мыши, так и узкочерепные полевки действуют со значительно большей скоростью и гораздо эффективнее, чем крысы или хомячки, повреждая и умерщвляя добычу серией укусов, так же как это делают бурозубки.
Согласно классификации охотничьего поведения мелких грызунов [6] можно предположить, что не только всеядные хомячки Кэмпбелла, зерноядные полевые мыши, но и зеленоядные узкочерепные полевки обладают прогрессивными чертами охотничьего стереотипа в виде захвата добычи лапами. Охотничий стереотип у этих видов более примитивный, чем у специализированных хищных грызунов, но обладает чертами высокой специфичности и может позволить расширить спектр пищевых ресурсов путем активной охоты на насекомых.
Исследования поддержаны Российским научным фондом (грант № 14-14-00603).
Библиографическая ссылка
Левенец Я.В., Пантелеева С.Н. СРАВНИТЕЛЬНЫЕ ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ ОХОТНИЧЬЕГО ПОВЕДЕНИЯ МЕЛКИХ МЛЕКОПИТАЮЩИХ // Современные проблемы науки и образования. 2016. № 6. ;URL: https://science-education.ru/ru/article/view?id=25813 (дата обращения: 19.06.2026).