



Органическое вещество – единственный источник энергии для почвы и формирования ее плодородия. Уровень почвенного плодородия определяется не только величиной запаса органического вещества, но и скоростью его минерализации. Микроорганизмы используют органический материал в качестве пищи и энергии [1].
Панцирные клещи, или орибатиды – одна из доминирующих по численности и биомассе групп почвообитающих беспозвоночных [5]. В настоящее время известно свыше 7000 видов. Панцирные клещи обладают высокой численностью, видовым разнообразием, достаточно эврибионтны, но отчетливо реагируют на экологические сдвиги в почвенной среде [3]. В численности панцирных клещей в микростациях под различными сельскохозяйственными культурами заметна закономерность увеличения общей численности в микростациях с растительностью и ее уменьшение в микростациях, лишенных растительности [6, 7, 8].
Достаточно специфично выглядит ситуация плотности населения панцирных клещей орибатид в естественных биогеоценозах. Анализ исследований, проведенных в естественных биотопах, показал, что в основном доминирующей группой здесь являются панцирные клещи, тогда как в агроценозах их численность снижается и возрастает численность акароидных, а иногда гамазовых клещей [2].
Цель данной работы: провести сравнительный анализ количественных показателей плотности населения панцирных клещей орибатид в естественных биотопах и агроценозах юга Тюменской области.
Материалы и методы исследования
Исследования проводились в 2005–2007 гг. с мая по сентябрь на полях учебно-опытного хозяйства Государственного аграрного университета Северного Зауралья. Почва на опытном поле – чернозем выщелоченный маломощный тяжелосуглинистый пылевато-иловый на карбонатном покровном суглинке. Растительность на территории учхоза представлена березовыми и березово-осиновыми лесами колочного, реже паркового типа, характерными для лесостепи Зауралья. Рельеф – слабоволнистая равнина с блюдцеобразными западинами с едва заметным уклоном на северо-восток. Отбор почвенных проб проводился по общепринятой методике [4]. Для взятия проб использовали бур объемом 125 см3 в десятикратной повторности на глубину до 15 см. Пробы отбирали в зернопаровом севообороте под овсом, однолетними травами (горохоовсяная смесь) и пшеницей, в пяти вариантах: 1) контроль; 2) внесение минеральных удобрений под 40 ц/га запланированного урожая; 3) внесение минеральных удобрений под 60 ц/га запланированного урожая; 4) внесение минеральных удобрений и добавление подкормки под 40 ц/га запланированного урожая; 5) внесение минеральных удобрений и добавление подкормки под 60 ц/га запланированного урожая. Вносимые на опытном поле удобрения: аммиачная селитра и диаммофос. Для подкормки вносили 30 кг азота в фазу кущения. На опытном поле проводились следующие обработки: ранневесеннее боронование; 2 культивации; посев сеялкой СКП – 2,1; прикатывание, отвальная вспашка (глубина 30 см). Отбор почвенных проб проводился также на целине (участок, не подвергавшийся никаким обработкам и расположенный в центре агроценоза) и в березово-осиновом колке (который находился в 50 м от агроценоза).
Методика обработки почвенных проб включала следующие этапы:
- извлечение микроартропод из почвенных проб – проводили при помощи термоэклектора Берлезе-Тульгрена. Для подсушивания пробы использовали электролампу 40 вт. Экспозиция длилась 5–7 дней при температуре 300–400 С;
- фиксация – традиционно используется этиловый спирт высоких концентраций (70–80 %), при этом рекомендуется добавлять глицерин (1–2 %), который сохраняет материал в случае случайного высыхания спирта в пробирке в процессе хранения;
- хранение – традиционно хранение в 70–80 % этиловом спирте, иногда рекомендуют более высокую концентрацию (96 %);
- обработка перед заливкой в препарат – особи переносятся из спирта в 5–10 % КОН, где по ходу просветления из темноокрашенных они превращаются в красные, а затем в прозрачно-розовые или коричневатые, липиды на поверхности тела исчезают;
- заливка в препарат – чистые и, если необходимо, просветленные экземпляры могут быть помещены в каплю консервирующей среды на предметном стекле.
Результаты исследования и их обсуждение
Выявляя влияние минеральных удобрений на динамику численности орибатид, следует отметить, что плотность населения панцирных клещей была наиболее существенной на участке, где вносили минеральные удобрения под 40 ц/га запланированного урожая (табл. 1).
Сравнив участок с внесением минеральных удобрений и добавлением подкормки под 40 ц/га) с контрольным и участками с внесением удобрений под 40 ц/га и под 60 ц/га, выделим, что на полях, где вносили только минеральные удобрения и не добавляли подкормку, плотность населения орибатид была существенно выше, чем на поле, где вносили минеральные удобрения и добавляли подкормку. Однако если сравнить с контрольным участком, то четко видно, что количество клещей на поле, где вносили минеральные удобрения и добавляли подкормку под 40 ц/га, существенно выше контроля (табл. 1).
Также необходимо заметить, что даже самые низкие по численности количественные показатели орибатид удобряемых участков превосходят по плотности контрольные участки примерно на 65 % (табл. 1).
Таблица 1
Плотность населения орибатид в агроценозах на участках с внесением различных доз минеральных удобрений (2005 г., 2006 г., 2007 г.) (экз./м2) (n=210)
|
Зернопаровой севооборот (овес, однолетние травы, пшеница) |
Участок |
||||
|
Контроль |
Удобрения под 40 ц/га |
Удобрения под 60 ц/га |
Удобрения и подкормка под 40 ц/га |
Удобрения и подкормка под 60 ц/га |
|
|
Среднее |
898,4 |
1683,7 |
1472,5 |
1374,0 |
1483,2 |
Изучая сезонные колебания численности орибатид в березово-осиновом колке, отметим (рис. 1), что плотность населения панцирных клещей была наиболее значительной весной (май) и ближе к осени (вторая половина августа).
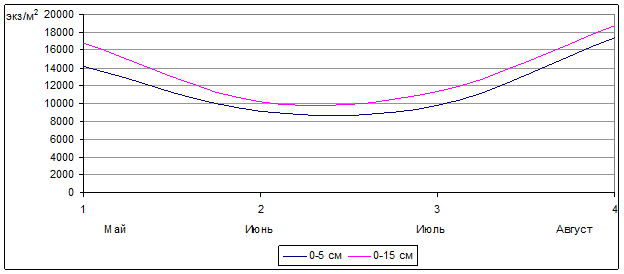
Рис. 1. Сезонная динамика численности орибатид в березово-осиновом колке на глубине почвы 0–5 см и 0–15 см (2005 г., 2006 г., 2007 г.) (экз./м2) (n=30)
Самые низкие показатели численности орибатид были отмечены в начале лета (июнь), возможно объяснение этого заключалось в недостатке пищевых ресурсов. Для перегнивания лесной подстилки (травы, листьев, и т.д.), являющейся основным источником пищи для панцирных клещей, необходимы определенные условия, прежде всего она должна достаточно прогреться и пройти первоначальную «обработку» микроорганизмами, уже после чего станет доступной для использования в пищу орибатидами. Возможно, данным процессом объясняется повышение численности панцирных клещей в июле и еще большее ее увеличение во второй половине августа, когда к перегнивающей прошлогодней подстилке добавляется постепенно разлагающийся корневой опад, травянистой и древесной растительности этого года (рис. 1).
Рассматривая сезонную динамику численности орибатид на целинном участке в течение трех лет (2005–2007 гг.), стоит выделить (рис. 2), что плотность населения панцирных клещей весной (май) была достаточно значительной (2667,3 экз./м2). В начале лета (июнь) показатели численности увеличиваются примерно в два раза, хотя в следующем месяце наблюдался небольшой спад. Ближе к осени количество панцирных клещей начинает резко увеличиваться, достигая 12053,3 экз./м2. Данное колебание численности орибатид, возможно, объясняется наличием достаточного количества корма, а также воздействием таких факторов, как температура и влажность почвы. Весной происходит отрождение новых особей орибатид и подъем из более глубоких слоев почвы к поверхностному, быстрее прогреваемому слою взрослых орибатид.
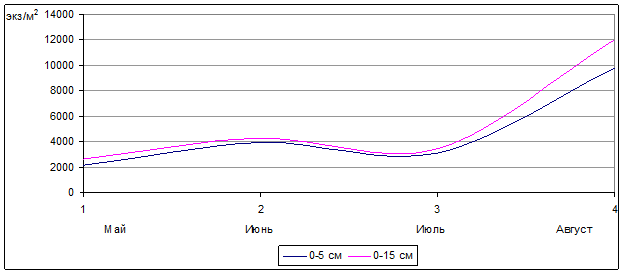
Рис.2. Сезонная динамика численности орибатид на целине на глубине почвы 0–5 см и 0–15 см (2005 г., 2006 г., 2007 г.) (экз./м2) (n=30)
Примерно в середине июня численность орибатид достигает своего первого пика (4315,3 экз./м2), характеризуемого большим количеством клещей разных возрастов. Ближе к июлю численность орибатид несколько сокращается (примерно на 1/4), возможно, это явление объяснялось нехваткой пищи. Однако результаты обработки почвенных проб, взятых во второй половине августа, свидетельствуют о том, что отмирающие и постепенно разлагающиеся остатки травянистой растительности создают кормовую базу клещей достаточной для питания. В связи с чем средняя численность панцирных клещей во второй половине августа достигала наивысших показателей за сезон (рис. 2).
При сравнении численности панцирных клещей, зафиксированных на целинном участке (5632,3 экз./м2) и в березово-осиновом колке (14266,8 экз./м2), с агроценозом, прежде всего, отметим, что плотность населения орибатид на данных участках (целина, березово-осиновый колок) (табл. 2) превосходила самые значительные по численности участки агроценоза (пшеничное поле 1556,4 экз/м2) – от 3 до 9 раз (табл. 3).
Таблица 2
Плотность населения орибатид в березово-осиновом колке и на целине (2005 г., 2006 г., 2007 г.) (экз./м2) (n=120)
|
Плотность населения орибатид |
||
|
Березово-осиновый колок |
Целина |
|
|
Среднее |
14266,8 |
5632,3 |
Также при сравнении этих участков можно пронаблюдать процесс изменения численности клещей в двух исследуемых слоях почвы (0-5 и 0-15 см). На участках, не подвергавшихся активному антропогенному воздействию (целина и березово-осиновый колок), численность орибатид как в поверхностном (0-5 см), так и в более глубоких слоях (0-15 см) практически синхронно изменялась в течение сезона (рис. 1,2), а на обрабатываемых участках в течение сезона плотность населения панцирных клещей в исследуемых слоях почвы имела существенные отличия.
Таблица 3
Плотность населения орибатид в агроценозах (2005 г., 2006 г., 2007 г.) (экз./м2) (n=350)
|
Возделываемая с/х культура |
Плотность населения орибатид |
|||
|
2005 г |
2006 г |
2007 г |
Среднее |
|
|
Овес |
1146,5±38,17 |
444,1±17,61 |
2325,2±88,53 |
1305,3 |
|
Однолетние травы |
1727,9±56,28 |
1017,0±43,74 |
1442,2±47,69 |
1395,7 |
|
Пшеница |
1121,7±48,91 |
1016,7±39,65 |
2530,8±104,18 |
1556,4 |
Возможно данное явление находит свое объяснение во вмешательстве человека в процессы жизнедеятельности клещей, идущие в почве, а именно – воздействие минеральных удобрений и механические обработки почвы, создающих то благоприятные условия существования для орибатид, то, наоборот, создающих для них стрессовые ситуации. Изменение этих условий среды обитания орибатид будет сопровождаться миграциями из более глубоких слоев к поверхностному и наоборот.
Сравнив целинный участок и березово-осиновый колок (рис. 1, 2), отметим, что численность панцирных клещей на обоих участках достигала значительных показателей. Однако березово-осиновый колок превышал по численности целинный участок примерно в 2,6 раза. Это, вероятно, объясняется более подходящими для орибатид микроклиматическими условиями березово-осинового колка, а также более богатой кормовой базой.
Выводы:
- количественные показатели плотности населения орибатид достигали наибольших значений в агроценозе на участке с внесением минеральных удобрений под 40 ц/га запланированного урожая (1683,7 зкз./м2);
- наибольшей плотности населения панцирные клещи (орибатиды) достигали под пшеницей (1556,4 экз/м2);
- плотность населения орибатид в исследуемом березово-осиновом колке (14266,8 экз./м2) и на целине (5632,3 экз./м2) превосходит самые значительные по количественным показателям участки агроценоза (пшеничное поле 1556,4 экз/м2) от 3 до 9 раз.
Библиографическая ссылка
Либерман Е.Л., Козлов С.А. СРАВНИТЕЛЬНЫЙ АНАЛИЗ ПЛОТНОСТИ НАСЕЛЕНИЯ ОРИБАТИД В ЕСТЕСТВЕННЫХ БИОТОПАХ И АГРОЦЕНОЗАХ ЮГА ТЮМЕНСКОЙ ОБЛАСТИ // Современные проблемы науки и образования. 2016. № 6. ;URL: https://science-education.ru/ru/article/view?id=25745 (дата обращения: 28.07.2026).