



Южный Урал и примыкающие к нему территории, которые можно условно обозначить как Приуралье, характеризуются большим разнообразием природных условий. Это определяется расположением региона на стыке крупных физико-географических единиц, детерминирующих интерградацию биотических комплексов разного генезиса, нахождением рассматриваемой территории в пределах двух природно-климатических зон (лесостепной и степной), а также разной степенью антропогенной нагрузки на экосистемы.
В итоге, на Южном Урале сформировались специфичные биотические комплексы, структурные особенности которых меняются не только в широтном или долготном направлении, но и в зависимости от рельефа местности, условий увлажнения, форм и интенсивности антропогенных влияний. В связи с этим анализ габитуса сообществ разных групп организмов представляет собой и вполне самостоятельную ценность, как один из аспектов изучения регионального биоразнообразия и в качестве инструмента исследования закономерностей варьирования характеристик среды региона.
Объектом нашего исследования были сообщества древоразрушающих грибов, существующие в березовых лесах Южного Урала. Березовые леса распространены во всех частях региона и занимают более 11 % общей лесопокрытой площади. Широкая встречаемость березняков в разных условиях делают их удобным объектом для сравнительного анализа компонентов их экосистем. Древоразрушающие грибы являются неотъемлемой частью любых лесных экосистем, играя решающую роль в функционировании системы редуцентов, производящей деструкцию древесной мортмассы; они также выступают в качестве регуляторов древостоев, вызывая стволовые и корневые гнили ослабленных экземпляров деревьев. Многими исследованиями [1, 5, 7 и др.] доказано, что структурные характеристики микоценозов древоразрушающих грибов могут использоваться в качестве вполне верных, хотя и косвенных индикаторов состояния лесных экосистем. Предмет наших исследований – варьирование габитуса ксиломикокомплексов березняков в зависимости от особенностей среды. В данном случае под габитусом мы понимаем облик сообществ, т.е. его основные измеряемые характеристики, такие как видовое разнообразие, систематическую, экологическую и функциональную структуру. Применение понятия габитус оправданно в том плане, что в природных условиях мы анализируем сообщества, опираясь на представленность плодовых тел тех или иных видов грибов в конкретных условиях, и наблюдаемая картина зависит от ряда факторов – сезон наблюдения, сохранность плодовых тел разных видов, профессиональный уровень исследователя и др. [11].
Материалы и методы
Сбор материалов о габитусе ксиломикокомплексов березняков проводился в 1994-2016 гг. Объект исследования – сообщества базидиальных ксилотрофных грибов (преимущественно афиллофороидных). Исследования проводились методом сплошного учета базидиом грибов на маршрутах с последующей идентификацией образцов с использованием специализированной русскоязычной и иностранной литературы [2, 3, 14-17]. Виды и надвидовые таксоны приведены в соответствие с системой грибов, представленной на Интернет-ресурсе «Index Fungorum» (дата обращения 15 октября 2016 г.).
При анализе габитуса сообществ использовались методические подходы, разработанные В.А.Мухиным [5], М.А.Сафоновым [6, 7].
Исследованиями были охвачены березняки разных районов Оренбургской области, а также Кугарчинского, Куюргазинского, Зиянчуринского, Зилаирского районов Республики Башкортостан. В общей сложности было обследовано 40 местообитаний; собрано и определено более 2300 образцов.
Результаты и обсуждение
В результате проведенных исследований в формационной микобиоте березняков Южного Урала выявлено 137 видов древоразрушающих грибов, относящихся к 68 родам и 28 семействам отдела Basidiomycota. Выявленные виды составляют 44,6 % от числа видов, выявленных в регионе [9]. Наиболее многочисленные рода Hyphoderma (7 видов), Hyphodontia, Trametes (по 6 видов), Peniophora, Polyporus, Postia (по 5 видов).
Видовое разнообразие изученных микоценозов варьирует в достаточно широких пределах – от 3 до 43. Это обусловлено рядом причин – разными площадями изученных березняков, отличиями в возрасте и состоянии древостоев. Большее значение для оценки габитусов сообществ имеет показатель видового богатства, а также соотношение между видовым разнообразием и численностью плодовых тел. Видовое богатство в изученных локалитетах отличалось значительным постоянством – в большинстве случаев оно составляло 4,4–5,0 видов на гектар. Лишь в отдельных искусственных березняках оно достигало 11 или снижалось до 1,7 вида на гектар. Соотношение между числом видов и количеством плодовых тел в изученных микоценозах также варьирует в широких пределах – от 0,13 до 0,83.
Важной чертой каждой формационной микобиоты являются специфичные виды, не встречающиеся в лесах других формаций – чаще всего по причинам субстратной специализации [8]. К числу видов, специфичных для березняков региона, относятся Anomoloma myceliosum (Peck) Niemelä & K.H. Larss., Climacodon septentrionalis (Fr.) P.Karst., Crustomyces subabruptus (Bourd. & Galz.) Julich, Inonotus obliquus (Ach. ex Pers.) Pilat, Irpex murashkinskyi (Burt) Kotir. & Saaren., Junghuhnia pseudozilingiana (Parmasto) Ryvarden, Lenzites betulinus (L.) Fr., Mycoacia aurea (Fr.)J.Erikss. & Ryvarden, Piptoporus betulinus (Bull.) P. Karst., Postia guttulata (Sacc.) Julich, Skeletocutis alutacea (J. Lowe) Jean Keller, Tyromyces fumidiceps G.F. Atk. , T.kmetii (Bres.) Bondartsev & Singer [12, 13]. При этом за исключением Piptoporus betulinus все эти виды малочисленны и представлены отдельными находками в березняках разных частей региона [4]. Наличие и участие в микоценозах специфичных видов – важная черта габитуса, определяющая ценность данного сообщества, так как высокая доля специфичных видов – показатель устойчивости сообщества во времени, показатель его сформированности.
Еще один важный показатель – соотношение сапротрофных и фитопатогенных видов в локальных микоценозах. Этот показатель позволяет прогнозировать дальнейшее развитие лесной экосистемы и самого микоценоза, так как высокая доля фитопатогенных видов свидетельствует о существенных перестройках в структуре древостоя, что, впоследствии, отразится и на судьбе микоценоза. В изученных микоценозах представленность фитопатогенных видов варьирует в широких пределах – от 0 до 16,7 % (по численности плодовых тел – от 18,5 до 71,5 %).
Как видно из приведенных данных, большинство показателей микоценозов варьирует в широких пределах, что позволяет предположить, что варьирование габитусов изученных сообществ происходит под влиянием более чем одного фактора среды. В частности, из приведенной карты (рис.1) видно, что видовое разнообразие микоценозов не связано с широтной зональностью, хотя наличие значимых связей видового состава позволяет предположить существование видовых комплексов приуроченных к березнякам лесостепной и степной зоны.
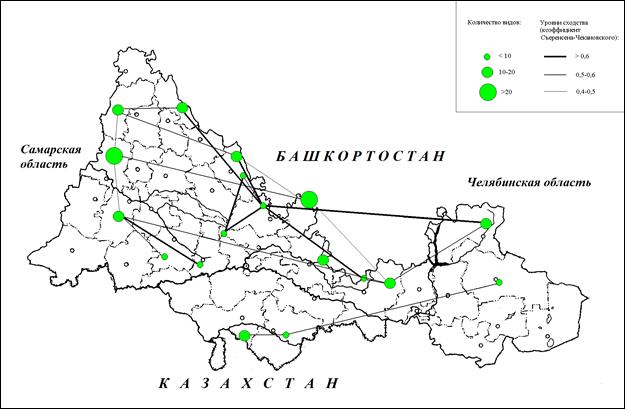
Рис. 1. Видовое разнообразие и сходство видового состава изученных ксиломикокомплексов березняков Южного Урала и Приуралья
Исходя из предположения, что на распределение видов древоразрушающих грибов и габитус их комплексов влияют условия увлажнения и температурный режим местообитаний, зависящие, в свою очередь, от ряда факторов (положение в рельефе, в той или иной природно-климатической зоне и т.п.), изученные микоценозы были объединены в несколько групп: микокомплексы березняков предгорий и низкогорий; микоценозы лесостепной зоны; подзоны настоящих степей; подзоны южных (сухих) степей; пойменных биотопов; Национального парка «Бузулукский бор». Комплексы грибов Бузулукского бора рассматриваются отдельно, так как этот реликтовый сосновый лесной массив является форпостом лесостепной биоты, вдающимся далеко на юг в пределы степной зоны, что накладывает определенный отпечаток на его биоту [10]. Выделенные группы присущи разным типам местообитаний, которые представляют собой серию биотопов, условия которых варьируют в комплексном экологическом градиенте.
Кластерный анализ сходства видового состава групп микокомплексов показал, что наиболее сходны между собой микокомплексы березняков, произрастающих в Бузулукском бору (1) и в подзоне разнотравно-злаковых степей (2) (рис. 2). Значительное сходство существует и между комплексами видов лесостепных березняков (3) и березняков предгорий и низкогорий (4). Наиболее обособлен комплекс видов, описанный в пойменных березняках (6).
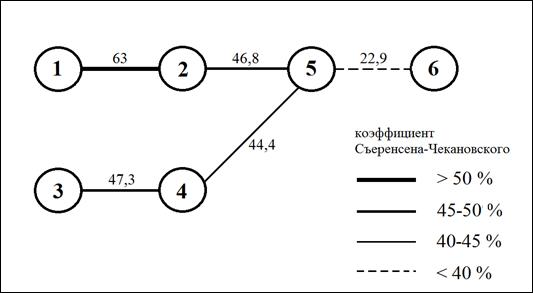
Рис. 2. Сходство видового состава групп микокомплексов березняков
Сравнение между собой выделенных групп микоценозов показало ряд отличий в их габитусе (таблица).
Средняя изменчивость показателей габитуса микокомплексов березняков в экологическом градиенте
|
Биотопическая приуроченность березняков |
число видов, шт |
видовое богатство, шт/га |
доля специфичных видов, % |
доля видов – фитопатогенов, % |
Концентрация доминирования
|
|
предгорья и низкогорья |
17,5 |
4,5 |
14,5 |
12,0 |
0,220 |
|
лесостепная зона |
15,75 |
4,4 |
15,7 |
11,0 |
0,246 |
|
степная зона (подзона настоящих степей) |
9,0 |
3,7 |
15,2 |
14,9 |
0,241 |
|
степная зона (подзона сухих, злаковых степей) |
13,0 |
4,0 |
5,0 |
10,5 |
0,149 |
|
пойма |
6,5 |
4,0 |
0 |
20,0 |
0,212 |
|
Бузулукский бор |
31 |
3,2 |
3,0 |
2,0 |
0,129 |
Так, при определенной выравненности видового богатства локальных микоценозов, о чем упоминалось выше, доля специфичных видов заметно выше в условиях северной части региона. Доля видов-фитопатогенов в сообществах снижается от березняков пойменных биотопов к березнякам Бузулукского бора. Низкая доля фитопатогенов в березняках национального парка объясняется высоким видовым разнообразием этого локального микокомплекса, что снижает показатель относительной значимости этих видов. Анализ концентрации доминирования, оцененной посредством индекса Симпсона, показывает, что максимальных значений она достигает в лесостепных, предгорных условиях и в березняках, произрастающих в подзоне настоящих степей. Минимальные показатели отмечены в сообществах березняков Бузулукского бора и в подзоне сухих степей.
Таким образом, можно сделать вывод, что ксиломикокомплексы березовых лесов Южного Урала обладают общими чертами, обусловленными во многом специфичной микобиотой. Габитус комплексов при этом существенно варьирует в комплексном экологическом градиенте, т.е. наблюдается не только закономерные изменения их физиономических характеристик при переходе от лесостепной к степной зоне, но и отличия, связанные с наличием азональных и интразональных условий, а также обусловленные положением березняков в рельефе.
Библиографическая ссылка
Сафонов М.А., Сафонова Т.И. ЗАКОНОМЕРНОСТИ ИЗМЕНЕНИЯ ГАБИТУСА МИКОЦЕНОЗОВ БЕРЕЗНЯКОВ В КОМПЛЕКСНОМ ЭКОЛОГИЧЕСКОМ ГРАДИЕНТЕ НА ЮЖНОМ УРАЛЕ // Современные проблемы науки и образования. 2016. № 5. ;URL: https://science-education.ru/ru/article/view?id=25452 (дата обращения: 29.06.2026).