



Одним из ключевых регионов при решении вопросов этногенеза народов Евразии является пограничная горная область, разделяющая Евразию на крупные регионы, – горные системы Алтая, Саян, Тянь-Шаня, Памира, Восточной Сибири. Эта горная область, тянущаяся вдоль степной полосы, на протяжении тысячелетий являлась ареной взаимодействия различных племен и народов двух крупнейших рас – европеоидной и монголоидной. Такие области интенсивных межэтнических контактов являются важным объектом популяционных исследований, поскольку горные системы служат не только барьерами на пути миграционных потоков, но и рефугиумами, сохраняющими следы генофондов разных эпох [3].
Горные системы Сибири сохранили следы древних популяций. Древнейшей (600–800 тыс. лет) стоянкой человека на территории Евразии считается стоянка Карама на Алтае. С похолоданием климата эта популяция Homo erectus переселилась или прекратила свое существование. Позднее (в период от 170–110 тыс. лет назад до 50–48 тыс. лет назад) здесь обитала другая линия – денисовский человек, названный так по имени Денисовой пещеры Алтая, в которой найдены его останки [8]. Денисовский человек во времени и пространстве перекрывается с неандертальцами, останки которых также обнаружены в горах Алтая (Денисова пещера, пещера Окладникова, Чагырская) и датируются возрастом 40–50 тыс. лет. Самые древние в Сибири представители человека анатомически современного вида – это человек из Усть-Ишима, на реке Иртыш (датировка – около 45 тыс. лет назад) и мальчик со стоянки Мальта близ Байкала (24 тыс. лет назад). В период неолита (VI–III тыс. до н.э.), благодаря «климатическому оптимуму», происходило активное заселение человеком самых отдаленных территорий Северной Евразии и закладывались основные этнокультурные общности. К периоду раннего металла сформировалась афанасьевская культура, появившаяся на рубеже IV–III тыс. до н.э. на Алтае и в Минусинской котловине. В первую четверть II тыс. до н.э. в южные районы Западной Сибири стали продвигаться племена андроновцев, в антропологическом облике которых обнаруживается и неоднородный европеоидный антропологический типа, и смешанный европеоидно-монголоидный антропологический тип [9].
В среднем бронзовом веке в Южной Сибири формируется окуневская культура (II тыс. до н.э.), антропологический состав которой включает монголоидную основу – автохтонный сибирский компонент, и европеоидный компонент, возможно западноевропейского происхождения. В конце II – начале I тыс. до н.э. в Южной Сибири сформировалась карасукская культура, сменившаяся тагарской культурой, в антропологическом типе которой преобладал европеоидный тип. В ранний железный век (VIII–VII века до н.э.) в Горном Алтае и Тыве возникают племенные союзы полукочевников – пазырыкский и уюкский.
Сменяющие друг друга культуры несли разные сочетания антропологических типов. Поэтому целью нашего исследования стало изучение влияния монголоидного (а в терминах генетики – восточно-евразийского) компонента на формирование современных генетических портретов народов Сибири и Центральной Азии – потомков носителей этих культур. Их изучение проведено с помощью SNP маркеров Y-хромосомы – наиболее эффективного инструмента популяционной генетики при анализе миграций. Исследованием охвачен обширный круг народов: Южной Сибири (алтайцев, тувинцев, хакасов, шорцев), Восточной Сибири (бурят, эвенков), Центральной Азии (казахов, киргизов, монголов).
Материалы и методы
Материалом исследования послужили образцы крови Биобанка населения Северной Евразии [4,6], собранные в ходе экспедиций Медико-генетического научного центра и Института общей генетик РАН с 2006 по 2015 год по единой технологии [5]. В Южной Сибири совместно с Кемеровским университетом собраны 733 образца крови шорцев, северных и южных алтайцев, хакасов. Совместно с учеными Республики Тыва – 257 образцов тувинцев и тоджинцев. В Восточной Сибири совместно с Забайкальским университетом – 867 образцов бурят и эвенков. Для сравнения использованы данные по SNP маркерам Y-хромосомы из базы данных Y-base, создаваемой в МГНЦ и ИОГен РАН под руководством проф. О.П. Балановского.
Для генетического анализа были отобраны наиболее информативные для народов Сибири и Центральной Азии гаплогруппы Y-хромосомы [1, 2]. Гаплогруппы для всех популяций были унифицированы до уровня следующих гаплогрупп: C(M130), C3(M217), C2(M38), C3c(M48), D(M174), E(M96), E1b1b1(M35), E1b1b1a(M78), G2a(P15), G2a1a(P18), G2a3b1(P303), J(M304), J1(M267), J2(M172), I(M170), I1(M253), I2a(P37), D2(P37.1), L(M20), L1(M27), L2(M317), L3(M357), N1(LLY22g), N1b(P43), N1c1(M178), O3(M122), O3a3(P201), O3a3a(M159), O3a3b(M7), O3a3c(M134), Q(M242), R(M207), R1b(M343), R1a1a(M198), R1b1b2(M269), R1b1b1(M73), T(M70) (классификация, предложенная в [Karafet et al., 2008] в модификации 2009 года, сделанной компанией Family Tree DNA). Анализ SNP маркеров проведен на Real Time ПЦР амплификаторе ABI 7900 флюоресцентным сканированием методом TaqMan (Applied Biosystems).
Результаты
Выявлен обширный спектр гаплогрупп (вариантов) Y-хромосомы, указывающий на большое генетическое разнообразие генофонда. Анализ главных компонент частот гаплогрупп у народов Сибири, Центральной Азии и других регионов Евразии был проведен в предыдущих работах [1, 2]. Но большой новый массив данных позволяет точнее оценить и степень сходства генофондов, и роль отдельных гаплогрупп. При анализе главных компонент выделяются «маркерные» или «сигнальные» гаплогруппы, определяющие разные пласты в генофонде коренного населения с условными названиями – «сибирский компонент»: N1c, N1b, Q; «западно-евразийский»: R1a, R1b, I; «центрально-азиатский» компонент: варианты гаплогрупп C, D и O. Наличие в генофонде народов гаплогрупп С, D и О рассматривается как свидетельство об экспансии монгольских племен [2].
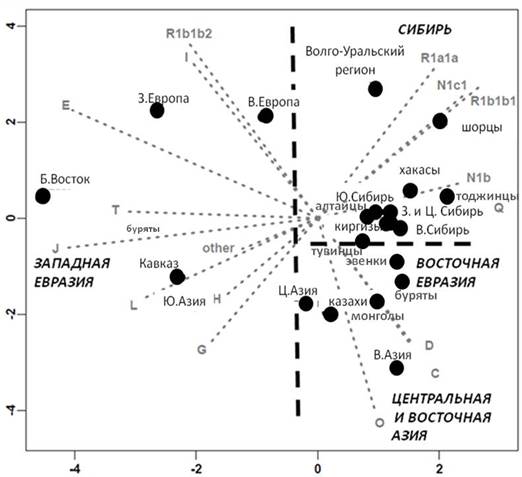
Рис.1. Положение изученных народов среди Евразийских генофондов в пространстве первой и второй главных компонент изменчивости гаплогрупп Y-хромосомы
Анализ распределения «маркерных» гаплогрупп в генофондах народов Южной, Восточной Сибири и Центральной Азии выявляет пестрый спектр гаплогрупп, который у степных популяций (казахов, монголов) значительно шире [10] и включает как восточно-евразийские гаплогруппы (C, D, O), так и западно-евразийские (R1b, I, J, G, E), а также варианты гаплогрупп N1c, N1b и Q, специфичные для сибирских народов.
Подробный анализ полиморфизма Y хромосомы у тюркоязычного населения Сибири и Центральной Азии показал [1, 3], что для большинства народов Сибири характерно наличие доминирующей гаплогруппы, но народы, находящиеся на границе со степной полосой Евразии, характеризуются более разнообразным спектром гаплогрупп (Рис.2). В генофондах популяций, граничащих с «степным коридором», обнаруживается существенный (от 10 % до 70 % генофонда) восточно-евразийский вклад, который может быть связан с экспансией центральноазиатских народов и в особенности монгольских племен. Это народы Южной и Восточной Сибири: буряты, тувинцы, эвенки, южные алтайцы (алтай-кижи и теленгиты); Центральной Азии – казахи, киргизы (которые до монгольской экспансии имели преимущественно европеоидный антропологический облик). Тогда как в более северных популяциях – шорцев, северных алтайцев (кумандинцев, тубаларов, челканцев), хакасов, тоджинцев – «восточно-евразийские гаплогруппы» не выявлены и преобладают «сибирские» и «западно-евразийские гаплогруппы».
Наиболее южные народности Южной Сибири - алтай-кижи и теленгиты - сочетают в своем генофонде практически весь спектр гаплогрупп, представленный у народов Сибири: причем одинаков вклад восточно-евразийских (С, D, O) и «сибирских» (N1c, N1b и Q) вариантов. Своеобразие генофонда тувинцев - при том же вкладе гаплогруппы С - проявляется в высоких частотах «сибирской» гаплогруппа Q [11] и центрально-азиатской гаплогруппы O (приближающейся к ее частоте в генофондах монголов и казахов).
Киргизы, генофонд которых формировался в Южной Сибири, сохранили «сибирские» варианты гаплогрупп (N1c, N1b и Q) [1, 2], но на более поздних этапах этногенеза подверглись мощному влиянию со стороны монгольских племен: варианты гаплогруппы С составляют у киргизов Тянь-Шаня больше трети генофонда, но у киргизов Памира, лучше сохранивших исходный генофонд, – лишь пятую часть генофонда. На восток «монгольское» генетическое влияние простирается до Тихого океана: оно проявляется и в Восточной Сибири [1, 2], и на Дальнем Востоке [7]. Однако гаплотипическое разнообразие гаплогруппы C максимально у монголов (около 60 %), указывая на центр их происхождения именно в Монголии. У прибайкальских бурят Иркутской области обнаружены наиболее высокие частоты субварианта гаплогруппы С-М407 (более 80 %), распространенного у монголов, калмыков и казахов и практически отсутствующего в генофондах народов Южной и Восточной Сибири. Исключение составляют тувинцы – этот вариант составляет у них 10 % генофонда. Забайкальские буряты (хоринские) существенно отличаются от бурят Иркутской области и Республики Бурятия. Их генофонд на 80 % состоит из одного субварианта гаплогруппы N1c1, характерного исключительно для бурят-хоринцев и монголов. Поскольку 80 % гаплотипов относятся к одному основателю, то можно считать столь высокую частоту этого варианта результатом резкого увеличения численности после прохождения популяцией «бутылочного горлышка». Забайкальские эвенки также несут весомый отпечаток экспансии монгольских племен: более половины их генофонда представлено вариантами гаплогруппы C.
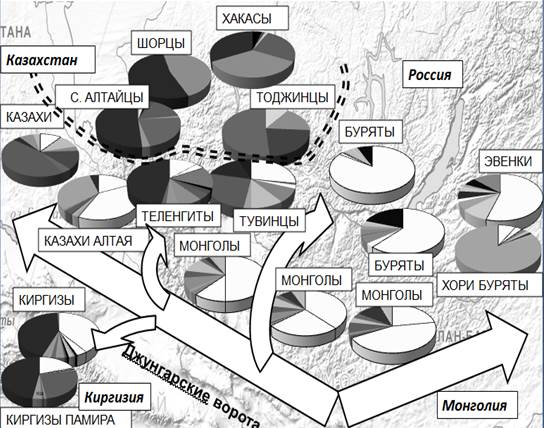
Рис. 2. Спектры гаплогрупп Y-хромосомы у народов Сибири и Центральной Азии
Примечание: белым цветом отмечены сектора гаплогруппы С Y-хромосомы, маркирующей монгольскую экспансию; черными секторами отмечена гаплогруппа R1a; стрелками обозначены основные миграционные потоки монгольских племен по данным Y-хромосомы.
Итоги исследования полиморфизма Y-хромосомы указывают, что поздние этапы формирования генофондов народов Южной и Восточной Сибири связаны с постепенным нарастанием восточно-евразийского компонента, проявляющегося в накоплении у современного населения Сибири «центрально-азиатских» вариантов гаплогруппы С. Широкая экспансия монгольских племен наиболее отразилась на генофондах народов, расселенных вдоль основной транспортной артерии кочевых племен – степной полосы Евразии. В западном направлении наибольший монгольский след прослеживается у тувинцев и южных алтайцев. В северо-восточном направлении максимальных частот гаплогруппа С достигает в генофонде бурят Иркутской области. Однако анализ гаплотипического разнообразия указывает, что источником миграций носителей этой гаплогруппы был ареал современной Монголии.
Библиографическая ссылка
Агджоян А.Т., Балаганская О.А., Схаляхо Р.А., Дамба Л.Д., Балановский О.П., Жабагин М.К., Юсупов Ю.М., Сабитов Ж.М., Богунов Ю.В., Балаганский А.Г., Султанова Г.Д., Долинина Д.О., Падюкова А.Д., Маркина Н.В., Букин А.Г., Лавряшина М.Б., Балановская Е.В. МОНГОЛЬСКИЙ СЛЕД В ГЕНОФОНДЕ НАРОДОВ ВДОЛЬ СТЕПНОЙ ПОЛОСЫ ЕВРАЗИИ // Современные проблемы науки и образования. 2016. № 4. ;URL: https://science-education.ru/ru/article/view?id=24910 (дата обращения: 01.07.2026).