



Оценка современного состояния биологического разнообразия лесных фитоценозов и их ресурсов играет важную роль в формировании как фундаментальных основ изучения и сохранения растительных ресурсов, так и прикладных аспектов их рационального использования в хозяйственно-экономических интересах страны. На основе оценки характера изменчивости и популяционной структуры можно наметить общие пути сохранения генофонда вида в регионе на популяционной основе, отобрать локальные популяций для сохранения и воспроизводства вида. Однако популяционный подход остается наименее разработанным в области сохранения биоразнообразия растений [7]. Изучение генофондов ресурсных видов растений с использованием молекулярных маркеров ДНК основано на анализе количественных характеристик генетического разнообразия популяций [3]. Для характеристики генетического разнообразия гетерогенных природных популяций растений необходим молекулярно-генетический анализ их генофондов с использованием, как минимум, двух типов высоко полиморфных молекулярных маркеров [2]. Эти маркеры должны давать возможность анализа полиморфизма различных структурных элементов генома. Так, ISSR-маркеры выявляют полиморфизм участков ДНК, заключенных между тандемно повторяющимися элементами – микросателлитами [10], и приобрели большую популярность для изучения генетического разнообразия ресурсных травянистых [1] и древесных видов растений [8]. Другой тип молекулярных маркеров (SNP-маркеры) выявляют нуклеотидный полиморфизм в локусах структурных генов. В совокупности два типа молекулярных маркеров позволяют проанализировать большую часть генома изучаемого вида, дать разностороннюю характеристику изучаемых генофондов и выявить их специфические особенности. Большую актуальность приобретает изучение генофондов ресурсных видов растений, занимающих обширные ареалы и имеющих хозяйственное значение, таких как виды рода Larix Mill., которые являются самыми распространенными древесными видами растений России и играют большую водоохранную и почвозащитную роль, особенно в северных и горных лесах. На Урале род Larix представлен западной расой лиственницы сибирской или л. Сукачева (Larix sukaczewii Dyl.). В связи с этим изучение генофондов популяций L. sibirica Северного и Среднего Урала на основе анализа полиморфизма двух типов ДНК-маркеров и разработка методики оценки состояния генофондов этого вида являются актуальной задачей для сохранения популяций лесных древесных видов, продуктивных и устойчивых к действию различных факторов среды.
Таким образом, цель данного исследования – дать характеристику и оценку состояния генофондов популяций западной расы лиственницы сибирской Северного и Среднего Урала на основании анализа полиморфизма двух типов молекулярных маркеров.
Материал и методы исследования
В качестве объектов исследований избраны девять природных популяций западной расы лиственницы сибирской, произрастающих на Среднем и Северном Урале, восемь из которых располагаются в Пермском крае: в государственном заповеднике «Вишерский» (Ls1, Ls2), около г. Красновишерск (Ls3); в Чердынском (Ls4), в Гаинском (Ls5), в Добрянском (Ls7), в Суксунском (Ls8) районах, а также две популяции находятся в Свердловской области: вблизи г. Качканар (Ls6) и пос. Билимбай (Ls9).
Для проведения молекулярно-генетического анализа в каждой из популяций собрана хвоя – индивидуально с 28–30 деревьев. Для выделения ДНК использовали методику С. Роджерса [9] с небольшими модификациями. Для полимеразной цепной реакции ISSR-методом реакционная смесь объемом 25 мкл содержала: 2 единицы Taq-полимеразы; 2,5 мкл 10х буфера + MgCl2 («Силекс М», Россия); 25 пМ праймера («Синтол», Россия); 0,25 мM dNTP («Fermentas», Литва); 5 мкл ДНК. Амплификацию ДНК проводили по стандартной для ISSR-метода программе с пятью ISSR-праймерами: М3 – (AC)8CT, Х10 – (AGC)6C, Х11 – (AGC)6G, ISSR-8 – (GAG)6C, CR-215 – (CA)6GT. Продукты амплификации разделяли электрофорезом в 1,7 % агарозном геле в 1х ТВЕ буфере, окрашивали бромистым этидием. Фотографирование и определения длины фрагментов ДНК проводили с помощью системы гель-документации GelDoc и программы Quantity One («Bio-Rad», USA). Для выявления SNP-маркеров в геноме L. sibirica отобраны три локуса потенциально адаптивно-значимых генов: 4CL1-363, sSPcDFD040B03103-274, ABA-WDS, амплификацию и секвенирование последовательностей данных локусов проводили в соответствии с рекомендациями, приведенными в литературных источниках [5].
Компьютерный анализ полиморфизма ДНК проведен с помощью общепринятых компьютерных программ POPGENE 1.31 и специализированного макроса GenAlEx6 для MS-Excel с определением доли (Р95) полиморфных локусов, абсолютного (na) числа аллелей, эффективного (ne) числа аллелей, ожидаемой (HE) гетерозиготности, информационного (I) Индекса Шеннона. Для выявления структуры внутрипопуляционного разнообразия применяли показатели внутрипопуляционного разнообразия (μ) и доли редких морф (h) [4]. Редактирование секвенированных последовательностей проводили в программе Sequence Scannerv1.0, местоположение полиморфных позиций определяли посредством выравнивания послeдоватeльностeй в компьютерной программе BioEdit v7.2.5. В программе DNASP v5 были реконструированы гаплотипы и на основе сравнения их нуклеотидных последовательностей рассчитаны общее гаплотипическое разнообразие (Hd) и нуклеотидное разнообразие (π), оценивающее среднее число парных различий на сайт. Оценка состояния генофондов популяций проведена в соответствии с методикой С. В. Боронниковой [2]. Выявлeниe спeцифичeских особeнностeй гeнофондов было провeдeно по модифицированной для дикорастущих дрeвeсных видов растeний мeтодикe подсчeта коэффициeнта гeнeтичeской оригинальности – КГО [6].
Результаты и их обсуждение
ISSR-маркеры в зависимости от праймера у L. sibirica выявляли от 17 до 34 ISSR-маркеров и всего в девяти популяциях было детектировано 123 ISSR-маркера, из которых 117 были полиморфными, то есть в совокупности изученные популяции характеризуются высокой долей полиморфных локусов (Р95=0,951). В отдельных популяциях этот показатель варьировал в пределах от 0,741 в популяции Ls7 до 0,871 в популяции Ls9 (табл. 1). Другой из основных показателей генетической изменчивости – ожидаемая гетерозиготность (HE). Она в суммарной выборке составила 0,202. Этот показатель выше в популяции Ls9 (HE=0,246), а самое низкое его значение (HE=0,171) отмечено в популяции Ls7. Для анализа генетического разнообразия на популяционном уровне был применен индекс разнообразия Шеннона. Он также выявил наибольшее разнообразие в популяции Ls9 (I=0,369), а наименьшее (I=0,258) – в популяции Ls7 (табл. 1). В дополнение к показателям, полученным на основании полиморфизма ISSR-маркеров, проведена характеристика генетического разнообразия изученных популяций на основе анализа гаплотипического разнообразия трех локусов потенциально адаптивно-значимых генов L. sibirica. Наибольшим гаплотипическим разнообразием характеризуется популяция Ls9 (Hd=0,911), а самое низкое значение этого показателя (Hd=0,636) выявлено в популяции Ls8. (табл. 1).
Большое значение для характеристики генофондов имеют показатели внутрипопуляционного разнообразия, анализ которого с применением показателя μ установил, что из 9 изученных популяций большей равномерностью распределения частот аллелей характеризуются популяции Ls5 и Ls9 (μ=1,747 и 1,738 соответственно), а наименьшей (μ=1,626) – популяция Ls7 (табл. 2). Показатель доля редких морф (h) оценивает структуру внутрипопуляционного разнообразия. По мнению Л. А. Животовского (1980), чем меньше значения h порогового 0.3, тем более сбалансированной структурой разнообразия характеризуются популяции. У всех изученных популяций показатель h имеет значения меньше 0,3 (табл. 1), наиболее сбалансированной структурой разнообразия характеризуется популяция Ls5 (h = 0,113), а наименее (h=0,187) популяция Ls7 (табл. 1). Для сравнения данных о внутрипопуляционном разнообразии, полученных с помощью двух типов ДНК-маркеров, проведен расчет показателя h на основании полиморфизма локуса 4CL1-363. В качестве морф рассматривалось число гаплотипов в популяции. В результате получены аналогичные ISSR-маркерам закономерности в распределении и структуре внутрипопуляционного разнообразия изученных популяций L. sibirica. Наиболее сбалансированной структурой нуклеотидного разнообразия характеризуются популяции Ls5 и Ls2 (h=0,078 и 0,075 соответственно), а наименее сбалансированной по данному типу маркеров (h=0,258) оказалась популяция Ls8 (табл. 1).
Для характеристики своеобразия генофондов большое значение имеет число уникальных аллелей (Un). Уникальные аллели, присутствующие только в одной из популяций и встречающиеся с заметной частотой (более 1 %) характеризуют уникальность генофонда на популяционном уровне. С использованием ISSR-метода анализа полиморфизма ДНК в популяциях Ls3, Ls5, Ls9 выявлено по одному уникальному, а в популяции Ls6 – 2 уникальных ISSR-маркера. Информативными для выявления уникальных аллелей были и SNP-маркеры. Уникальные SNPs обнаружены в восьми популяциях L. sibirica: в популяции Ls3 найдены 2 уникальных SNPs, а в популяции Ls7 – 5, в остальных выявлено по 1 уникальному SNP-маркеру. В среднем с наибольшей частотой SNPs встречались в популяции Ls5 (0,450), а с меньшей (0,130) – в популяции Ls4 (табл. 1).
Установленные при молекулярно-генетическом анализе параметры генетического разнообразия L. sibirica разделены на четыре группы (табл. 1). К первой (1) группе относятся «Основные показатели генетического разнообразия», вторую группу (2) параметров генетического разнообразия составляют показатели внутрипопуляционного разнообразия, третья группа (3) характеризует генетическую структуру и дифференциацию популяций (табл. 1). Важной группой параметров генетического разнообразия является четвертая (4), так как характеризует «специфику» генофонда. Высокие значения КГО популяции свидетельствуют о повышенном присутствии редких для региона исследований аллелей и соответственно специфичности ее генофонда, а популяции с минимальным значением КГО имеют минимальные частоты и число редких аллелей. Такие популяции характеризуются наиболее типичным или базовым генофондом [6]. При характеристике специфичности генофондов необходимо учитывать не только значения КГО, но также присутствие и число уникальных аллелей в популяции, так как наличие таких аллелей может указывать на особенности структуры генетического разнообразия популяций. Наибольшие значения КГО имеет популяция Ls5 (КГО=1,171), кроме того в ней обнаружены уникальные аллели, выявленные с помощью как ISSR-, так и SNP-маркеров (табл. 1). Эта популяция является самой западной и располагается на большом расстоянии от других изученных популяций, вероятно в связи с этим в ней сосредоточены нетипичные для региона исследований аллели, что может быть обусловлено историей расселения вида в регионе и формированием его ареала из различных популяционных систем. Вторым по величине КГО и, соответственно, нетипичными аллелями обладает девятая популяция L. sibirica (КГО=0,939). Она расположена в предгорной области восточного макро-склона вблизи границы Среднего и Южного Урала и является самой южной из изученных популяций. В ней также обнаружены уникальные аллели, выявленные с помощью обоих типов ДНК-маркеров.
Таблица 1
Оценка состояния генофондов популяций L. sibirica
|
Популя- ция |
1. Основные показатели генетического разнообразия |
2. Внутрипопуля- ционное разнообразие |
3. Генетическая структура и дифференциация |
4. Специфика генофондов |
|||||
|
P95 |
HE |
I |
μ |
Hd |
hISSR |
hSNP |
UnISSR/SNP |
КГО |
|
|
Ls1 |
0,772 |
0,172 |
0,271 |
1,639 |
0,872 |
0,181 |
0,154 |
0/1 |
0,935 |
|
Ls2 |
0,806 |
0,182 |
0,287 |
1,664 |
0,871 |
0,168 |
0,075 |
0/1 |
0,925 |
|
Ls3 |
0,806 |
0,189 |
0,298 |
1,698 |
0,887 |
0,151 |
0,111 |
1/2 |
0,760 |
|
Ls4 |
0,821 |
0,225 |
0,313 |
1,703 |
0,857 |
0,148 |
0,117 |
0/0 |
0,587 |
|
Ls5 |
0,871 |
0,223 |
0,338 |
1,774 |
0,836 |
0,113 |
0,078 |
1/1 |
1,171 |
|
Ls6 |
0,816 |
0,213 |
0,328 |
1,714 |
0,859 |
0,143 |
0,091 |
2/1 |
0,776 |
|
Ls7 |
0,741 |
0,168 |
0,258 |
1,626 |
0,883 |
0,187 |
0,160 |
0/5 |
0,703 |
|
Ls8 |
0,768 |
0,200 |
0,305 |
1,664 |
0,636 |
0,168 |
0,258 |
0/1 |
0,930 |
|
Ls9 |
0,872 |
0,242 |
0,369 |
1,738 |
0,911 |
0,131 |
0,126 |
1/1 |
0,939 |
Примечание: P95 – доля полиморфных локусов; HE – ожидаемая гетерозиготность; I – информационный индекс Шеннона; μ – показатель внутрипопуляционного разнообразия; Hd – общее гаплотипическое разнообразие; hISSR – доля редких морф, рассчитанная на основании ISSR-маркирования; hSNP – доля редких морф, рассчитанная на основании SNP-маркирования; UnISSR/SNP – число уникальных аллелей, выявленных с помощью ISSR- и SNP-маркеров; КГО – коэффициент генетической оригинальности.
Наименьшее значение КГО имеет четвертая популяция (КГО=0,587), располагающаяся на севере Пермского края почти в центре района среднетаежных лесов, занимающих здесь огромные территории с непрерывным ареалом и практически не затронутые человеческой деятельностью (табл. 1). Вероятно, вследствие этого популяция Ls4 характеризуется наиболее типичным генофондом. Уникальные маркеры в данной популяции не обнаружены. Таким образом, для ранжирования генофондов по степени специфичности предлагаются следующие интервалы: с КГО от 0,580 до 0,930 – типичный; с КГО от 0,930 и выше – специфичный, но при этом необходимо также учитывать наличие уникальных аллелей. Так, в популяции Ls7 с КГО=0,703 выявлено наибольшее число (5) уникальных (то есть специфичных) аллелей, выявленных с помощью SNP-маркеров, поэтому генофонд данной популяции может быть отнесен к специфичным (табл. 1).
Для оценки состояния генофондов популяций все избранные показатели генетического разнообразия переведены в разработанную на примере изученных природных популяций L. sibirica шкалу оценки состояния генофондов. На основании данной шкалы установлено, что в высокой степени удовлетворительное состояние (Ia) характерно для генофондов популяций Ls5 и Ls9 (табл. 2); в средней степени удовлетворительном (Iб) состоянии находятся генофонды шести популяций L. sibirica (табл. 2), а в одной популяции наблюдается тенденция к обеднению генофонда (Ls7), его состояние оценено как средняя степень обеднения – IIa (табл. 2, рисунок).
Таблица 2
Шкала оценки состояния генофондов популяций L. sibirica
|
Состояние генофонда |
P95 |
HE |
I |
μ |
Hd |
hISSR/SNP |
Un1 |
КГО |
||
|
I Удовлетворительное |
>0,750 |
>0,170 |
>0,300 |
>1,650 |
>0,750 |
<0,200 |
>3 |
>0,900 |
||
|
Iа |
высокая степень |
>0,820 |
>0,220 |
>0,350 |
>1,700 |
>0,870 |
<0,150 |
>1,100 |
||
|
Iб |
средняя степень |
0,750-0,820 |
0,170-0,220 |
0,300-0,350 |
1,650-1,700 |
0,750-0,870 |
0,200-0,150 |
0,900-1,100 |
||
|
II Обеднение генофонда |
0,550-0,750 |
0,100-0,170 |
0,200-0,300 |
1,500-1,650 |
0,550-0,750 |
0,300-0,200 |
1-2 |
0,500-0,900 |
||
|
IIа |
средняя степень |
0,650-0,750 |
0,130-0,170 |
0,250-0,300 |
1,550-1,650 |
0,650-0,750 |
0,250-0,200 |
0,700-0,900 |
||
|
IIб |
сильная степень |
0,550-0,650 |
0,100-0,130 |
0,200-0,250 |
1,500-1,550 |
0,550-0,650 |
0,300-0,250 |
0,500-0,700 |
||
|
III Деградация генофонда |
<0,550 |
<0,100 |
<0,250 |
<1,500 |
<0,550 |
>0,300 |
<1 |
<0,500 |
||
|
IIIа |
средняя степень |
0,450-0,550 |
0,050-0,100 |
0,200-0,250 |
1,300-1,500 |
0,450-0,550 |
0,350-0,300 |
0,400-0,500 |
||
|
IIIб |
сильная степень |
<0,450 |
<0,050 |
<0,200 |
<1,500 |
<0,450 |
>0,350 |
<0,400 |
||
Примечание: P95 – доля полиморфных локусов; HE – ожидаемая гетерозиготность; I – информационный индекс Шеннона; Hd – общее гаплотипическое разнообразие; μ – показатель внутрипопуляционного разнообразия hISSR/SNP – доля редких морф, рассчитанная на основании ISSR- и SNP-маркирования; Un1 – общее число уникальных аллелей, выявленных с помощью ISSR- и SNP-маркеров; КГО – коэффициент генетической оригинальности.
С целью сохранения генофонда ценного ресурсного вида растений L. sibirica рекомендуется отбирать как популяции с типичными (базовыми) генофондами, так и популяции, обладающие специфическими особенностями генофондов, являющиеся резервом генетической изменчивости.
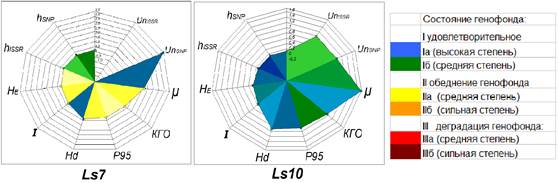
Оценка состояния генофондов на примере двух популяций L. sibirica; P95, HE, I, Hd, μ, hISSR, hSNP, UnISSR, UnSNP, КГО – показатели генетического разнообразия; справа – шкала оценки состояния генофондов
Для отбора в качестве объектов сохранения могут быть рекомендованы популяции со специфическими генофондами, обладающие высоким уровнем генетического разнообразия. Например, такие как популяции Ls5 и Ls10, характеризующиеся высокими значениями КГО и наибольшими показателями генетического разнообразия. Кроме того, популяция Ls7, для которой характерно в целом обедненное состояние генофонда, имеет наибольшее число уникальных SNPs-маркеров и высокий показатель гаплотипического разнообразия, поэтому может быть рекомендована для отбора в качестве резерва генетической изменчивости на нуклеотидном уровне при сохранении генофонда вида (рисунок). Для сохранения базовых генофондов лиственницы сибирской предлагается популяция Ls4, для которой характерно в целом в средней степени удовлетворительное состояние генофонда, отсутствие уникальных маркеров и сбалансированная генетическая структура. Так же типичным генофондом обладает популяция Ls2. Данная популяция характеризуется высоким уровнем генетической изменчивости и внутрипопуляционного разнообразия, доля редких морф (hISSR=0,168; hSNP=0,075) свидетельствует о практически ненарушенной генетической структуре. Она находится на территории заповедника «Вишерский» на высоте около 800 метров над уровнем моря и поэтому может быть использована для сохранения генофонда L. sibirica в горах.
Заключение
Таким образом, на основании данных о генетическом разнообразии, полученных с помощью двух типов молекулярных маркеров, установлено, что в удовлетворительном состоянии находятся генофонды восьми изученных популяций, а в одной популяции (Ls7) отмечены признаки обеднения генофонда. Наибольшие значения коэффициента генетической оригинальности (КГО) как интегрального показателя популяционного разнообразия установлены в популяциях Ls5 и Ls10. Для отбора в качестве объектов сохранения генофондов рекомендуются популяции с типичными и специфическими генофондами. Изучение генетической изменчивости природных популяций древесных растений и оценка состояния их генофондов могут быть использованы для составления генетически обоснованных программ по сохранению, восстановлению и рациональному использованию лесных генетических ресурсов.
Работа выполнена при частичной финансовой поддержке программы «УМНИК» Фонда содействия развития малых форм предприятий в научно-технической сфере (2013–2015 гг.).
Библиографическая ссылка
Нечаева Ю.С., Жуланов А.А., Красильников В.П., Боронникова С.В. ОЦЕНКА СОСТОЯНИЯ ГЕНОФОНДОВ ПОПУЛЯЦИЙ ЗАПАДНОЙ РАСЫ ЛИСТВЕННИЦЫ СИБИРСКОЙ LARIX SIBIRICA LEDEB. (L. SUKACZEWII) НА СРЕДНЕМ И СЕВЕРНОМ УРАЛЕ // Современные проблемы науки и образования. 2016. № 3. ;URL: https://science-education.ru/ru/article/view?id=24520 (дата обращения: 02.07.2026).