



В настоящее время известны так назывемые С3-путь и С4-путь фиксации СО2, фотосинтез по типу толстянковых (САМ-метаболизм) и фотодыхание (гликолатный цикл). С3-путь фотосинтеза (цикл Кальвина) – основной и присущий всем растениям, химизм расшифрован М. Кальвином и его сотрудниками в 1946–1956 гг. Первичным продуктом ассимиляции углерода является фосфоглицериновая кислота (СH2O(P)-CHOH-COOH). В ее молекуле содержится 3 атома углерода. Поэтому такой способ связывания углекислоты называли С3-путем фотосинтеза.
В результате дальнейших исследований был открыт еще один цикл фотосинтеза (1966), названный циклом М. Д. Хетча - К. Р. Слэка, или С4-циклом. Последнее название связано с тем, что в первичном продукте – оксалоацетате содержится в своей молекуле 4 атома углерода. К группе растений с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго, амарант, лебеда и многие другие виды Poaceae (мятликовые – злаки) и др. Для листьев С4-растений характерно анатомическое строение кранц-типа. Хлоропласты клеток обкладки содержат много зерен крахмала и не имеют гран. В клетках мезофилла хлоропласты имеют обычную гранальную структуру и так далее. Фотоситез по типу толстянковых (САМ-метаболизм) был обнаружен впервые у растений семейства Crassulaceae (толстянковых), и поэтому данный тип обмена углерода был назван метаболизмом кислот по типу толстянковых (Crassulaceae acid metabolism). Сокращено - САМ-метаболизм. Выявлено около 20000 видов растений, для которых характерен САМ-тип углеродного обмена.
В растительных клетках, содержащих хлоропласты, помимо вышеуказанных путей фотосинтеза, осуществляется также фотодыхание, т.е. активируемый светом процесс высвобождения СО2 и поглощения О2, который значительно отличается от «темнового» дыхания митохондрий. При этом первичным продуктом является гликолевая кислота, и поэтому путь получил название гликолатного.
В течение гликолатного цикла (фотодыхания) происходит потеря СО2, поглощенного в С3-цикле. В результате у С3-растений потери сухой массы составляют от 15 до 50 %. Было доказано, что фотодыхание осуществляется и у САМ-растений, но не так активно, как у С3- растений. Одназначного ответа на вопрос о наличии гликолатного цикла (фотодыхания) у С4-растений пока нет [2,3].
Из изложенного видно, что мировое обеспечение продовольствием может быть улучшено не только с помощью традиционной селекции и методов генной инженерии, но и освоения новых культур, принадлежащих в особенности к группе растений с С4, путем фотосинтеза как наиболее продуктивные.
Одним из них является амарант – однолетнее травянистое растение рода Amaranthus, семейства Амарантовые (Amaranthaceae). В мире известно 65 родов и 850 видов, распространенных в основном в тропических и субтропических областях земного шара. На территории Российской Федерации встречается 16–17 видов, из которых 12 видов распространены как культурные [6,11]. В условиях Кабардино-Балкарской Республики все изученные коллекционные образцы (78) различного географического происхождения (в том числе из Германии, Ямайки, Камеруна, Венгрии, Танзании, Узбекистана, Румынии и других стран) проходили полный вегетационный период с получением полноценных семян и высокой биомассы (900–3000 ц/га) с более высоким содержанием в белках незаменимых аминокислот, в том числе лизина, по сравнению с другими кормовыми и пищевыми растениями [11].
Установленные авторами закономерности жизнедеятельности амаранта и разработанные на этой основе способы получения высокого урожая семян и зеленой биомассы можно использовать не только для целей кормопроизводства и пищевой промышленности, но и как источник тепловой энергии. Нынешние разведанные и доступные энергетические ресурсы (нефть, газ, уголь, торф, горючие сланцы) – продукты разложения наземных и морских растений, микроорганизмов и животных, оцениваются в 1019-1020ккал, а годовое потребление энергии человечеством составляет более 1017ккал и ежегодно возрастает. Нетрудно подсчитать, что эти запасы могут закончиться в течение ХХI столетия [3]. Как отмечает автор, реальной и неисчерпаемой альтернативой существующим ныне видам энергии (помимо термоядерного синтеза) может служить только солнечная энергетика, которая основана на превращении энергии Солнца в электрическую (фотоэлементы, солнечные батареи). С другой стороны, в качестве топлива также эффективно могут использоваться и продукты фотосинтеза, как целлюлоза, интенсивно растущих видов древесных растений, кустарников и трав, в том числе, как отмечено выше, и биомасса амаранта.
Цель исследования – установить особенности изменения основных параметров фотосинтетической деятельности: площадь листьев, интенсивность и чистую продуктивность фотосинтеза, содержание пигментов и число хлоропластов в клетках листьев и их влияние на продуктивность растений при разных агроэкологических условиях выращивания амаранта.
Методы исследования
Опытные участки посевов амаранта располагались в предгорной зоне Кабардино-Балкарской Республики на территории ботанического сада КБГУ. Объектами исследований служили 78 образцов различного географического происхождения. В данной работе приводятся результаты, полученные в 1992–1994 гг. на примере одного из перспективных для КБР образцов – Амаранта метельчатого (A.cruentus) при оптимальных условиях проведения опытов по схеме:
1. Без внесения в почву удобрений (контроль),
2. N90 Р90 К90,
3. N45P65K30+ навоз 20 т/га.
Почвы – выщелоченные малогумусные среднемощные черноземы. Площадь делянки 120 м2, повторность опыта трехкратная. Густота посева семян 70х35 см.
Изучение мезоструктуры листьев проводили по методике [4]. Фотосинтетическую деятельность изучали, определяя площадь листьев методом высечек [7], интенсивность фотосинтеза (ИФ) – с помощью прибора газоанализатора в аппарате Варбурга, чистую продуктивность (ЧПФ) методом А. А. Ничипоровича и др. [8], содержание хлорофилла «а» и «б» в листьях с использованием спектрофотометра СФ-4. Результаты исследований обрабатывали методом математической статистики [1].
Результаты и обсуждение
Основным критерием оптимизации физиологических процессов и влияния агроэкологических факторов на растение является величина и качество урожая [9].
Наши исследования показали, что в варианте опыта без дополнительного внесения в почву основных элементов минерального питания (контроль) сырая надземная биомасса в среднем за 1992-1994 гг. достигает 830,5 ц/га, а воздушно-сухой вес семян – 15,09 ц/га. При внесении в почву азота, фосфора и калия по 90 кг действующего вещества на 1 гектар величина зеленой биомассы возрастает до 1345ц/га, а семян – до 19,16 ц/га, а при сочетании внесения в почву минеральных удобрений (N45P65K30) с органическими (навоз 20 т/га) соответственно биомасса до 1673 ц/га и семян до 22,41 ц/га при динамике роста амаранта в процессе онтогенеза в указанных условиях корневого питания [таблица 1]. Такая закономерность свидетельствует о высокой отзывчивости амаранта к плодородию почвы.
Продуктивность растений определяется за счет фотосинтетической деятельности растений в посевах [8]. В этой связи занимались изучением разных параметров фотосинтетического аппарата исследуемого образца амаранта [таблица 2]. Так, из данных таблицы видно, что внесение в почву минеральных и в особенности органических удобрений в сочетании с минеральными способствует увеличению площади листьев до 53,8 – 58,6 тыс.м2/га. При таких величинах площади листьев наблюдается увеличение интенсивности фотосинтеза до 46,5- 50,8 мг СО2/дм2 в фазах массовой бутонизации и цветения. Из данных таблицы также видно, что при улучшении условий корневого питания возрастает также величина чистой продуктивности фотосинтеза (ЧПФ) от 14,20 до 19,02 г/м2 за сутки. Приведенные данные свидетельствуют о том, что повышение площади листьев, мощности фотосинтетического аппарата, под действием элементов питания приводит к возрастанию как интенсивности фотосинтеза, так и чистой продуктивности данного процесса, уникального явления природы.
Таблица 1
Динамика роста амаранта метельчатого в процессе онтогенеза в зависимости от условий корневого питания в см (среднее за 1992–1994 гг.)
|
Варианты опыта |
От всходов |
||||
|
до образования 10 листа |
фазы массовой бутонизации |
фазы массового цветения |
фазы созревания семян |
Общая высота растений перед уборкой |
|
|
Без удобрений (контроль) |
19,7± 0,24 |
66,2±0,15 |
148,8±0,21 |
167,2±0,16 |
171,6±0,47 |
|
N90P90K90 |
26,6±0,61 |
98,5±0,42 |
224,7±0,51 |
247,6±0,34 |
256,2±0,28 |
|
N45P65K30 + навоз 20 т/га |
34,2±0,17 |
126,2±0,19 |
265,5±0,32 |
286,2±0,56 |
289,4±0,63 |
Среди многочисленных факторов, влияющих на интенсивность и продуктивность фотосинтеза, важное значение имеют число хлоропластов в клетках и на единицу площади листа, общее содержание хлорофилла («а»+ «в») в листьях [5]. С количественными характеристиками пластидного аппарата тесно связана фотосинтетическая «работоспособность», а, следовательно, и продуктивность растений. Вместе с тем именно на этом структурном уровне фотосинтетический аппарат растений обладает большими возможностями экологической пластичности, поскольку здесь существует целый спектр параметров, которые могут меняться в зависимости от условий среды. Выяснено, что в клетках опытных растений, выращенных в условиях минерального и особенно органо-минерального питания, увеличивается количество хлоропластов в клетке и на единицу площади листа, содержание общей суммы хлорофилла («а»+ «в») в листьях.
Таблица 2
Параметры фотосинтетического аппарата амаранта метельчатого при разных условиях корневого питания. Среднее за 1992–1994 гг.
|
Варианты опыта |
Интенсивность фотосинтеза(мгСО2/дм2×час) ЧПФ (г/м2× сутки) |
Площадь листьев , тыс м2/га |
Общее содержание хлорофилла «а» + «в», мг/дм2
|
Число хлоропластов на единицу площади листа среднего яруса, млн/см2 |
||||
|
ФЦ |
||||||||
|
Фаза массовой бутонизации (ФМБ) |
Фаза цветения (ФЦ) |
ФМБ |
ФЦ |
ФМБ |
ФЦ |
Палисад-ная ткань |
Губча-тая ткань |
|
|
Без удобрений (контроль) |
24,8 ±0,15 9,00± 0,17 |
26,4 ±0,13 11,02±0,19 |
34,5 |
36,2 |
2,00 |
2,32 |
1,545 |
1,475 |
|
N90P90K90 |
46,5±0,14 14,2±0,20 |
48,4 ±0,11 16,03±0,04 |
53,8 |
54,5 |
3,54 |
4,62 |
2,676 |
2,094 |
|
N45P65K30 + навоз 20 т/га |
48,7±0,12 18,7±0,11 |
50,08±0,61 19,02±0,07 |
56,4 |
58,6 |
3,86 |
4,98 |
3,264 |
2,856 |
По нашим данным, структура мезофилла и толщина листа мезофитного растения амаранта изменяются также в зависимости от образцов, условий корневого питания, расположения листьев по ярусам. Установлено, что у опытных растений воздействию различного минерального питания в основном подвергаются губчатая паренхима, клетки обкладки и толщина листа. Столбчатая паренхима изменяется реже (рисунок). Наибольшее увеличение указанных параметров, как было сказано выше, наблюдается при внесении минеральных и органо-минеральных удобрений. Таким образом, с внесением питательных веществ общая толщина листа, в том числе и мезофилла, увеличивается.
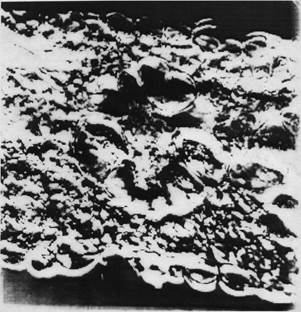
Анатомическое строение листа А. caudatus L. (кранц-типа) амаранта хвостатого К-27153, ув.10х40
Таким образом, можно сделать вывод, что элементы минерального и особенно органо-минерального питания, изменяя в лучшую сторону основные параметры фотосинтетического аппарата амаранта – С4-группы растений способствуют значительному повышению продуктивности растений, т.е. урожая зеленой биомассы и семян с 1 гектара посева. Увеличение продукционного процесса может происходить и за счет отсутствия у амаранта полуденной депрессии фотосинтеза и фотодыхания [2,3, 11].
Работа выполнена в свете научно-технической программы «Амарант», созданной в России при Санкт-Петербургском университете.
Библиографическая ссылка
Слонов Л.Х., Шугушева Л.Х., Слонов Т.Л. ФОТОСИНТЕТИЧЕСКИЙ АППАРАТ И ПРОДУКТИВНОСТЬ АМАРАНТА - С4 ГРУППЫ РАСТЕНИЙ // Современные проблемы науки и образования. 2016. № 2. ;URL: https://science-education.ru/ru/article/view?id=24191 (дата обращения: 07.07.2026).