



Световая кривая фотосинтеза - кривая, отражающая зависимость интенсивности фотосинтеза от плотности лучистого потокападающей фотосинтетическиактивной радиации. Световая кривая характеризует эффективность использования лучистой энергии на фотосинтез. Значение плотности светового потока, при котором наступает светонасыщение фотосинтеза, — величина непостоянная и зависит от экологических условий и видовых свойств растений [6].Напряженность любого внешнего фактора, выходящая за пределы нормы реакции генотипа, создает условия экологического стресса. Наиболее часто в наших условияхфактором экологического стресса для наземных растений является высокое содержаниесолей в почве, чтоприводит к расширению областей, имеющих крайне деградированный травянистый и почвенный покров.
Целью настоящего исследования являлосьизучение информативности световых кривых флуоресценции и фотосинтеза растений, произраставших в условиях различной степени засоленности и гумусированности почвы.
Материалы и методы исследования
Отбор материала, объектов исследования и измерение квантового выхода флуоресценции и фотосинтеза в условиях различной степени засоленности почвы проводили в Тарумовском районе Республики Дагестан на территории Качубейской биосферной станции. Для исследования зависимости флуоресцентных показателей растений от концентрации гумуса и степени засоленности почвы объекты исследования отбирались на площадках с предположительно разным уровнем засоления: участок №1 («бугристый солончак») - геоморфологическая обл. и район – приморская полоса Терско-Кумской низменности, слабонаклонная равнина на север, мезорельеф бугристый. Растительность – сарсазановые сообщества: сарсазан, солянка однолетняя, полынь, кермек, лебеда, амарант и др. Солончак типичный, глубокопрофильный (-21,9 – -23,7 н.у.м.). Координаты: шир. 440 31,4724 долг. 460 36,9549; участок №2 («лугово-болотная») - луговая слабо - или среднесолончаковая, слабогумусированная, среднесуглинистая (-21,7 – -24,0 н.у.м.). Координаты: шир. 440 30,7054; долг. 460 38,4856; участок №3 –«солончак типичный»глубокопрофильный с солончаками луговыми, глубокопрофильными, среднесуглинистыми (тяжелосуглинистые и глинистые), (-13,8 – -15,8 н.у.м.). Координаты: шир. 440 26,3222; долг. 460 26,0874.
Образцы почв отбирали по генетическим горизонтам на глубинах 0-10 см и 20-30 см на всех трех опытных площадках.
Для определения общей концентрации водорастворимых солей в почвах проводился анализ водной вытяжки по общепринятым методам [1]. Определяли плотный остаток, состав анионов (щелочность от нормальных карбонатов СО3-- и бикарбонатов НСО3-, анионы Cl-, SO4--) и состав катионов (Са2+, Mg2+, Na+).
Проводилось определение химического состава водных вытяжек почвы на степень засоленности, содержание гумуса(%)и концентрацию азота (мг/100г) в почвах с трех опытных площадок. Измерения параметров флуоресценции растений проводились на портативном хлорофилл - флуориметре анализаторе эффективности фотосинтеза MINI-PAM HeinzWalzGmbH. Математическую обработку материалов проводили с применением статистического пакета «Statistica 6».
Результаты и обсуждение
На глубине 20-30см содержание гумуса заметно меньше, чем на глубине 0-10 см (рис. 1 А).В бугристом солончаке почти в 4 раза (3,97), в лугово-болотной почве в 5 раз меньше (возможно из-за легковесности самого гумуса, очень влажно); в солончаке типичном почти в три раза (2.7). Это, возможно, зависит от плотности и увлажненности почвы. Концентрация азота в бугристом солончаке на глубине 0-10 см выше, чем на глубине 20-30см; в лугово-болотной почве выше в 2 раза; в солончаке типичном также выше в 2 раза (рис. 1 В).
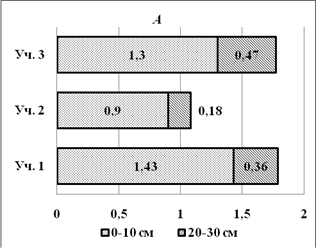
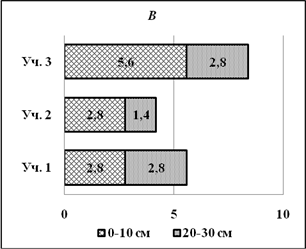
Рис. 1.Содержание гумуса (А) и азота (В) в почвах с различной солевой нагрузкой на глубине 0-10 и 20-30см.
На уровне 0-10 см концентрация азота в бугристом солончаке равна концентрации азота в лугово-болотной почве.В бугристом солончаке условия для разложения органических остатков наименее благоприятные (наибольшая засоленность). Высокая влажность в лугово-болотной почве и наибольшая отдаленность от моря опытной площадки № 3 (солончак типичный) способствует более успешному разложению органических остатковна поверхности почвы (0-10см).
На бугристом солончаке и лугово-болотной почве на глубине 0 – 10см плотный остаток практически на одном уровне, всолончаке типичном меньше в 16 раз (рис. 2). Т.о., светло-каштановая карбонатная почва с участка № 3 наименее засолена. На глубине 20-30 см засоленность почвы в бугристом солончаке в 2,6 раз выше, чем в лугово-болотной
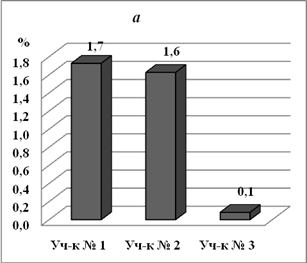
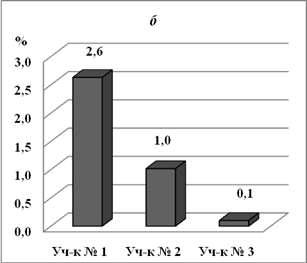
Рис. 2. Плотный остаток в почвах с опытных участков с различной степенью солевой нагрузки (%) на разных глубинах: а -0-10 см, б - 20-30 см.
почве и в 26 раз выше.чем на солончаке типичном. Т.е. в лугово-болотной почве на глубине 20-30 см засоленность ниже, чем на поверхности. Излишняя увлажненность лугово-болотной почвы, по-видимому, приводит к выпотному режиму засоления и поднятию солей на поверхность.
Естественные потери световой энергии при открытых реакционных центрах (F) в начале световой кривой (первый насыщающий световой импульс) минимальный у растений сбугристого солончака и максимальный с лугово-болотной почвы (рис. 3) (обратная корреляция с концентрацией гумуса и азота (рис. 2)). К концу световой кривой максимальный уровень F у растений с бугристого солончака, минимальный с солончака типичного. Т.к. растения на бугристом солончаке подвержены воздействию высокихконцентраций солей и более закалены к действию стресса другого рода, первый импульс растения выдерживают лучше и теряют световую энергию меньше, чем растения с менее засоленных участков. Но длительную атаку световыми импульсами растения выдерживают меньше и к восьмому импульсу истощаются и теряют световую энергию больше, что указывает на значительные фотоповреждения фотосинтетических систем.
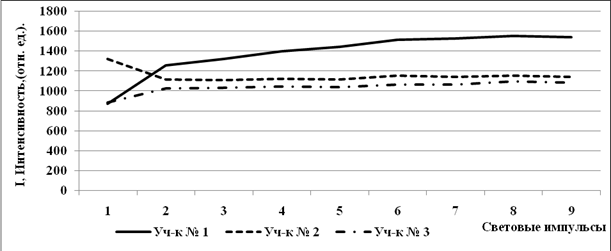 |
Рис. 3. Световые кривые квантового выхода флуоресценции (F)растений, произраставших на почвах с различной степенью солевой нагрузки.
Максимальная флуоресценция (Fm) при первом импульсе растений сбугристого солончака и с лугово-болотной почвы на одном уровне и выше, чем у объектов с солончака типичного (рис. 4).
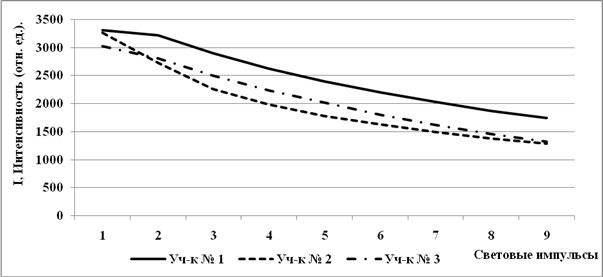
Рис. 4. Световые кривые квантового выхода максимальной флуоресценции (Fm) растений, произраставших на почвах с различной степенью солевой нагрузки.
К концу световых кривых показания максимальной флуоресценции
растенийс лугово-болотной почвы и солончака типичного на одном уровне и ниже, чем у растенийс бугристого солончака. У растений с бугристого солончака показания Fm на протяжении всей световой кривой держится выше двух других участков, что свидетельствует о значительных потерях световой энергии при световом насыщении и уже закрытых реакционных центрах.
При начальном насыщающем импульсе низкий выход Fm растенийсолончака типичного указывает на меньшие потери световой энергии и на то, что растения находятся в более благоприятных условиях произрастания по признаку засоленности (рис. 2). По всей световой кривой показания Fm ниже показаний с других участков,проходят у растений с лугово-болотной почвы. Наблюдается прямая корреляция показаний интенсивности максимальной флуоресценции с содержанием гумуса в почве (рис. 1 А) Снижение показаний максимальной флуоресценции у растений с лугово-болотной почвы свидетельствует о том, что идет смена флуоресцентных потерь на безизлучательные потери световой энергии (тепловая диссипация) и может рассматриваться как фотопротекторная реакция.
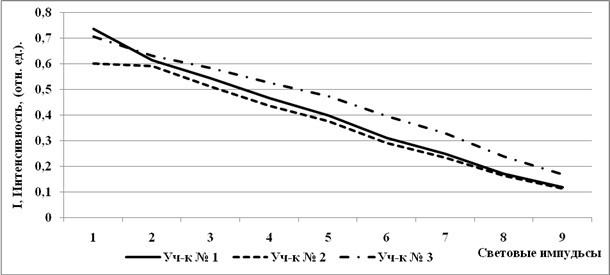 |
Минимальные значения квантового выхода фотосинтеза (Y) при первом световом импульсе наблюдаются у растений с лугово-болотной почвы, максимальные с бугристого солончака (рис. 5). Дальше световая кривая фотосинтеза выше других проходит у растений с солончака типичного, т.е. здесь наиболее высокий квантовый выход фотосинтеза и устойчивость растений к атакам световых импульсов.
Рис. 5. Световые кривые квантового выхода фотосинтеза (Y)растений, произраставших на почвах с различной степенью солевой нагрузки.
Световые кривые Y растений с лугово-болотной почвыи бугристого солончака проходят заметно ниже, что свидетельствует о низком уровне фотосинтеза в этих условиях при действии светового стресс-фактора. Наблюдается прямая корреляция интенсивности фотосинтеза с концентрацией азота в почве на глубине 0-10 см (рис. 1 В) и обратная корреляция с засоленностью (рис. 2). Выход фотосинтеза падает под действием всех факторов, ингибирующих реакционные центры фотосистемы II, при этом растет тепловая диссипация или показания флуоресценции.
Необходимо отметить, что различия в световых кривых фотосинтеза растений Кермек Мейера (галофит)с трех разных по засоленности и гумусированности опытных площадок, не такие существенные, как мы наблюдали в световых кривых флуоресценции.Это свидетельствует о поддержании растением уровня фотосинтеза на максимально возможном высоком уровне, независимо от условий произрастания, ввидуслишком высокой значимости процесса фотосинтеза для развития целого растения и перспективы дальнейшей передачи качественного генетического материала. Фотосинтез - процесс, обеспечивающий растительную клетку энергией,поэтому сохранение фотосинтетической активности в условияхфизиологического стресса во многом определяет устойчивость растения кнеблагоприятным факторам окружающей среды.
Заключение
Обнаруженные нами корреляционные зависимости в характере световых кривых флуоресценции F растений от общей засоленности, Fm от содержания гумуса и фотосинтеза Y от концентрации азота в почвах свидетельствует о присутствии заметной реакции оптических параметров растений на различия в состоянии почвы и позволяет проводить интегральную комплексную оценку состояния почвенной среды по флуоресцентной реакции высших растений на действие избыточного количества солей, гумусированность, концентрацию азота, суммарный эффект которых очень сложно выявить стандартными химическими методами контроля. Высокое временное разрешение, экспрессность, чувствительность и возможность автоматического измерения большого числа параметров флуоресценции растений в естественной природной среде позволяет рекомендовать применение метода в биомониторинге почвенной среды для своевременного обнаружения возникновения признаков аридизации на самых начальных этапах проявления.
Рецензенты:Асадуллаев З.М., д. б. н., профессор, директор института ФГБУН Горный ботанический сад ДНЦ РАН, г. Махачкала;
ГасановГ.Н., д.с. х.н., руководитель лаборатории биогеохимии,ФАНО РОССИИ, ФГБУН Прикаспийский институт биологических ресурсов Дагестанского научного центра РАН,г. Махачкала.
Библиографическая ссылка
Магомедова М.Х.-М., Маммаев А.Т., Алиева М.Ю., Пиняскина Е.В. ХАРАКТЕР СВЕТОВЫХ КРИВЫХ РАСТЕНИЙ, КАК ПОКАЗАТЕЛЬ СОСТОЯНИЯ ПОЧВЫ АРИДНЫХ ТЕРРИТОРИЙ // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/ru/article/view?id=23951 (дата обращения: 04.07.2026).