



В последнее время усиливается антропогенное загрязнение окружающей среды, связанное в первую очередь с поступлением тяжелых металлов в основные биогеохимические цепи. Изучение воздействия тяжелых металлов на растения привлекает в последнее время все большее внимание в связи с поступлением загрязнителей и в пищевые цепи [6]. Проблема усугубляется тем, что в отличие от других промышленных отходов ТМ, в том числе и соли стронция не разлагаются и уровень их содержания в среде со временем возрастает. Сведения о токсичности стронция для растений неоднозначны [4]. Исследования стрессовых реакций, вызванных действием солей стронция, имеют практическое и фундаментальное значение, поскольку связанно с изучением механизмов адаптации и устойчивости растений к тяжелым металлам.
Цель нашей работы - исследование действия тяжелых металлов на биометрические параметры пшеницы и флуоресцентные показатели фотосинтетической активности.
Методы и объекты исследования
В качестве объектов были использованы изогенные линии пшеницы Rht 9, Rht 13, Rht 17 сорта «Мироновская 808».
Семена пшеницы проращивали в чашках Петри на фильтровальной бумаге в вегетационной камере (23/21°C день/ночь; относительная влажность 70%; постоянная аэрация раствора). На 3 сутки после проклевывания проростки переносили в контейнеры с керамзитом. Семена были выращены в растворах хлорида стронция с концентрациями 1*10-4 Моль/л, 1*10-5 Моль/л, 1*10-6 Моль/л. Контрольные образцы были выращены на воде. Измерения проростков пшеницы производились на 5, 10 и 15 сутки после проклевывания. Учитывались темпы прорастания, общую всхожесть, прирост побегов, количество корней и длину главного корня. Количество повторностей 7-10. Статистическую обработку данных проводили с помощью пакетов стандартных программ Microsoft Excel 2007 и Statistica 6. Достоверность различий между парными значениями оценивали по t –критерию Стьюдента при р<0.05.
Параметры флуоресценции хлорофилла листьев измеряли с помощью флуориметра MINI-PAM (Pulse Amplitude Modulation) Yeinz Walz GmbH, (Германия) методом амплитудной импульсной модуляции света.
Перед измерением побеги были адаптированы к темноте в течение 10 мин. В ходе экспериментов регистрировали следующие параметры флуоресценции: F0 — интенсивность флуоресценции хлорофилла в адаптированных к темноте образцах при действии зондирующих импульсов возбуждающего света; Fm — интенсивность флуоресценции хлорофилла во время действия 0.8 с насыщающей вспышки света, восстанавливающей первичные хинонные акцепторы Qa до QА*; Y - эффективность процессов фотосинтеза; qP и qN - коэффициенты химического и нехимического фототушения флуоресценции, NPQ - параметр нехимического фототушения флуоресценции.
Для возбуждения флуоресценции хлорофилла используется импульсная модуляция красного света, источником которого служит «красный» светодиод (LED), максимум излучения при длине волны 650 нм. Продолжительность импульсов 3 мкс модуляционная частота 0,6 или 20 кГц. В так называемом режиме укороченной вспышки (burst-mode) вспышки продолжительностью 0,2 сек. чередуются с периодами темноты 0,8 сек. Свет светодиода проходит через фильтр (Balzers DT Cyan, special), с максимумом пропускания 650 нм и оставляющий небольшой участок спектра более 700 нм. Флуоресценция регистрируется с помощью трехслойного полупроводникового фотодиода (PIN-фотодиод) при длине волны более 700 нм, благодаря наличию длинноволнового фильтра (RG 9, Schott).
Интенсивность 0,15 мкмоль м-2с-1 ФАР; Источником насыщающих световых импульсов и постоянного актиничного освещения служит миниатюрная галогенная лампа 8 Вт /20 Вт (Bellaphot. Osram).
Результаты исследований и обсуждение
 Наши исследования показали, что во всех образцах соли стронция снижали всхожесть (рис.1). Наиболее подверженной к действию солей стронция оказалась высокорослая линия Rht 13. Всхожесть не превышала 50% при всех разведениях (см.рис.1). У линии Rht 9 при концентрации стронция 10-4 всхожесть семян совпадала с контрольной, при концентрациях 10-5 и 10-6 всхожесть составляла 80% от контрольной. У семян линии Rht 17 всхожесть при всех разведениях была стабильной и составляла »81%.
Наши исследования показали, что во всех образцах соли стронция снижали всхожесть (рис.1). Наиболее подверженной к действию солей стронция оказалась высокорослая линия Rht 13. Всхожесть не превышала 50% при всех разведениях (см.рис.1). У линии Rht 9 при концентрации стронция 10-4 всхожесть семян совпадала с контрольной, при концентрациях 10-5 и 10-6 всхожесть составляла 80% от контрольной. У семян линии Rht 17 всхожесть при всех разведениях была стабильной и составляла »81%.
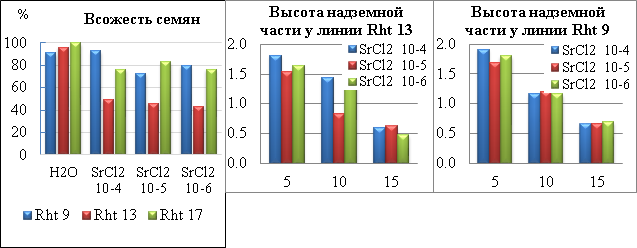 Одним из видимых симптомов действия стронция на изогенные линии пшеницы - нарушение роста и морфогенеза исследуемых растений. Мы провели морфометрические исследования проростков на 5, 10 и 15 сутки инкубации в растворах SrCl2. Отмечено, что на 5 сутки ростовые показатели превышали контрольные значения. Длина гипокотиля превышала контрольные показатели в 2 раза. У линий Rht 9 и Rht 17 на 5 сутки прирост главного корня по отношению к контролю был не более 20%. У линии Rht 13 на 5 сутки наблюдали прирост в длину как надземной, так и подземной частей. Причем показатели гипокотиля превышали контрольные на 80%, а длина главного корня больше контроля на 100-130%. Измерение длины надземной части у всех линий выращенных в различных концентрациях стронция показало прирост по отношению к контролю на 5 сутки и его дальнейшее постепенное ингибирование у Rht 9 и Rht 13 (рис. 1(б) и (в)). У низкорослой линии Rht 17 на 10 сутки наблюдали резкий скачок ростовых процессов как корневой, так и надземной части. Необходимо отметить, что концентрации, ингибирующие рост гипокотиля стимулировали рост корня в длину.
Одним из видимых симптомов действия стронция на изогенные линии пшеницы - нарушение роста и морфогенеза исследуемых растений. Мы провели морфометрические исследования проростков на 5, 10 и 15 сутки инкубации в растворах SrCl2. Отмечено, что на 5 сутки ростовые показатели превышали контрольные значения. Длина гипокотиля превышала контрольные показатели в 2 раза. У линий Rht 9 и Rht 17 на 5 сутки прирост главного корня по отношению к контролю был не более 20%. У линии Rht 13 на 5 сутки наблюдали прирост в длину как надземной, так и подземной частей. Причем показатели гипокотиля превышали контрольные на 80%, а длина главного корня больше контроля на 100-130%. Измерение длины надземной части у всех линий выращенных в различных концентрациях стронция показало прирост по отношению к контролю на 5 сутки и его дальнейшее постепенное ингибирование у Rht 9 и Rht 13 (рис. 1(б) и (в)). У низкорослой линии Rht 17 на 10 сутки наблюдали резкий скачок ростовых процессов как корневой, так и надземной части. Необходимо отметить, что концентрации, ингибирующие рост гипокотиля стимулировали рост корня в длину.
Концентрация стронция в растениях значительно превышает его содержание в почве и накапливается в больших количествах [3]. Содержание стронция в растениях, как и кальция, увеличивается на более поздних стадиях развития растений [2]. Характер накопления стронция в растениях близок к кальцию. Взаимодействие между этими элементами весьма сложно. Они могут конкурировать между собой, но стронций обычно не может заменить кальций в его биохимических функциях.
Исследуя действие солей стронция на проростки пшеницы разных линий мы работали в основном с биометрическими параметры этих растений. Так как биометрические показатели являются результатом изменений на молекулярном уровне, было исследовано действие металла на процессы фотосинтеза. Наиболее чувствительна к ионам металлов ФС II, активность которой оценивается на основании анализа кинетики флуоресценции хлорофилла. Изменения в присутствии Sr таких параметров флуоресценции хлорофилла, как максимальный (Fm) и минимальный (F0) выход флуоресценции (Y), переменная флуоресценция (Fυ), a также квантовая эффективность ФС II (Fυ/Fm), указывают на определенные нарушения в ФСА растений [5].
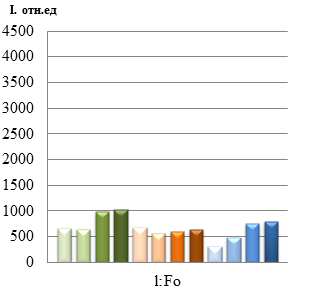
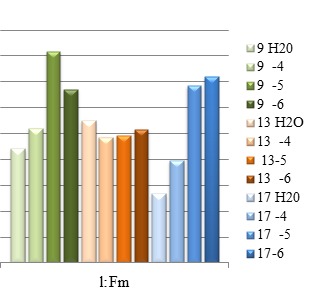
Рис.2 (а,б,в) Квантовые выходы флуоресценции (F) и максимальной флуоресценции (Fm) линий Rht 9,13,17 в зависимости от концентрации солей Sr
Полные данных о фотосинтетических реакциях можно получить из определения индукционной кинетики. При освещении зеленого листа, который был адаптирован к темноте (10 мин), интенсивность флуоресценции, как правило, претерпевает ряд немонотонных изменений (эффект Каутского). В результате исследований нами было показано, что квантовый выход флуоресценции (F) как и максимальной флуоресценции хлорофилла (Fm*) у всех линий пшеницы при концентрации Sr 10-6 моль/л совпадают с контрольными ( по воде) или являются максимальным (рис. 3 а,б,в). Снижение квантового выхода (F) у сорта Rht 13 указывает на фотоповреждение фотосинтезирующих систем.
Выход максимальный флуоресценции (Fm*), измеренный после полной темновой адаптации (когда все реакционные центры открыты и все первичные акцепторы электронов окислены) у линий Rht 9 и 17 максимален, при минимальном использовании энергии на фотохимические реакции и тепловое излучение. Уменьшение Fm* у линии Rht 13 (при всех разведениях) свидетельствует об увеличении безызлучательных потерь световой энергии -тепловая диссипация. Анализ кривых тушения флуоресценции (NPQ) показывает увеличение значений, приближаясь к контрольным (относительно воды), что свидетельствует об интенсификации процессов нефотохимического тушения, связанных с рассеиванием энергии возбуждения антенного хлорофилла в виде тепла. В целом, по рис. 2 видно, что наиболее чувствительной к Sr является линия Rht 13, даже слабая концентрация растворов металлов у которого оказывает негативное влияние на фотосинтетическую активность: уменьшение F и Fm*, рост qP и qN. Что же касается высокой концентрации Sr, то изучение индукционных кривых флуоресценции указывает на индукцию процессов нефотохимического тушения (см. рис.3 б), изменение скорости потока электронов по электронно-транспортной цепи (ETR), связанное с модификацией мембран хлоропластов, изменением состояния тилакоидов и общим ингибированием процессов фотосинтеза.
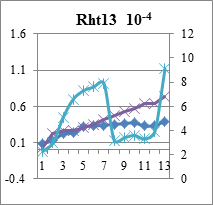
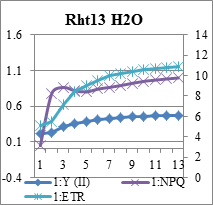

Рис.3. Индукционные кривые пшеницы линии Rht 13 - контроль (Н2О) и опыт (концентрации Sr10-4 и 10-6М). По левой вертикальной оси – интенсивность флуоресценции (I, отн. ед.), по правой вертикальной – скорость электронов по ETR (µmol electrons/(m2·s).
Концентрация 10-6 моль/л вызвала снижение фотосинтетической активности ниже исходного уровня. Нужно отметить, что индукционные параметры кинетических кривых при самой слабой концентрации у 10 дневных проростков пшеницы значительно отличались от 15 дневных: фиксировались резкие перепады величин qP, qN, ETR и Y, видимо к 15 дням проростки частично адаптировались, поскольку эффективность фотохимического преобразования энергии в ФСII (квантовый выход фотосинтеза) значительно выше (см. рис.4).
Анализируя эффективность фотохимического преобразования энергии в ФСII (квантовый выход фотосинтеза - Y),отметим, что наивысшее значение - 0,78 у сорта Rht 17 при концентрациях 10-5 и 10-6 моль/л, соответственно, превышая контрольные значения (Rht 17) или приравниваясь к ним (Rht 13). Несколько ниже значения у сорта Rht 9, причем если высокие концентрации стронция (10-4, 10-5) моль/л все-таки были выше контрольных, то концентрация 10-6 моль/л вызвала снижение фотосинтетической активности даже ниже исходного уровня. При изучении кинетики восстановления фотосинтетического аппарата наблюдалась та же картина: максимально быстро восстанавливались сорта Rht 17 (1мин.) и 13 (1,5 мин.), тогда как Rht 9 потребовался более продолжительный период времени до 4 минут.
Таким образом, в результате проведенной работы можно сделать следующие выводы: соли кадмия и стронция по-разному влияют на морфофизиологические показатели и фотосинтез растений. Степень ингибирования фотосинтеза во многом зависит от концентрации металла в субстрате. Высокие концентрации металлов влияют на ультраструктуру хлоропластов, замедляют скорость электронного транспорта, уменьшая содержание фотосинтетических пигментов и активность ферментов цикла Кальвина как за счет их прямого действия на отдельные реакции фотосинтеза, так и в результате опосредованного влияния на другие физиологические процессы.
Выводы
Инкубирование проростков пшеницы в растворах SrCl2 подавляло и всхожесть, и ростовые процессы во всех исследуемых концентрациях. При действии на проростки пшеницы SrCl2 ингибирующий эффект с течением времени усиливался. Исследованные концентрации металла снижали интенсивность фотосинтеза в период их воздействия. На основании анализа индукционных кривых флуоресценции хлорофилла всех сортов можно предположить, что на ранних стадиях токсическое действие Sr в исследованных концентрациях проявляется не только в подавлении электронного транспорта ФСII , но и в уменьшении степени энергизации фотосинтетических мембран.
Начальные процессы развития токсического действия Sr связаны, в первую очередь, со снижением эффективности световых реакций фотосинтеза, что и определяет в дальнейшем ухудшение продукционных свойств.
Библиографическая ссылка
Пиняскина А.В., Пиняскина Е.В., Гаджиева И.Х. ВЛИЯНИЕ СОЛЕЙ СТРОНЦИЯ НА БИОМЕТРИЧЕСКИЕ И ФЛУОРЕСЦЕНТНЫЕ ПОКАЗАТЕЛИ И ПШЕНИЦЫ // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/ru/article/view?id=23790 (дата обращения: 04.07.2026).