



В современной нейрофизиологии и нейрохимии особое внимание уделяется изучению механизмов пластичности нервной системы, с помощью которых обеспечиваются такие важнейшие функции, как обучение и память. Исследование молекулярных основ этих процессов необходимо как для понимания механизмов высших нервных функций, так и для поиска способов влияния на патологические состояния нервной системы. В связи с этим изучение механизмов действия факторов, влияющих на пластичность нервной системы, является одной из актуальных задач нейробиологии.
В качестве средств коррекции нарушенных функций мозга все чаще рассматривается особый класс веществ – регуляторные пептиды, которые являются важнейшими компонентами в функционировании основных систем организма: нервной, иммунной и эндокринной [1, 8]. Помимо широкого спектра фармакологических свойств, этот класс веществ обладает такими преимуществами, как полное отсутствие токсических и побочных влияний, гормональной активности, как правило, мягкий модуляторный характер действия [1, 8].
В Институте молекулярной генетики РАН под руководством академика Н.Ф. Мясоедова разработаны и синтезированы лекарственные средства на основе регуляторных пептидов: аналог адренокортикотропного гормона (АКТГ4-10) – семакс (Met-Glu-His-Phe-Pro-Gly-Pro) и производное тафцина – селанк (Thr-Lys-Pro-Arg-Pro-Gly-Pro). В клинических исследованиях показана их высокая эффективность при лечении ителлектуально-мнестических расстройств, а также терапевтическая роль при сосудистых заболеваниях головного мозга, в комплексной терапии при черепно-мозговых травмах [1, 3]. Однако результаты исследований последних лет не дают полного представления о спектре физиологической активности препаратов и о механизмах их нейротропного действия.
Известно, что первостепенная роль в формировании различных интегративных реакций организма, прежде всего, в мотивационно-эмоциональной сфере, отводится одному из важнейших элементов лимбической системы – гиппокампу. Участие данной структуры в развитии мотивационных состояний отчетливо проявляется в пищедобывательном поведении [5]. В частности, поле СА1 гиппокампа принимает непосредственное участие в формировании пространственной памяти животных. Гиппокампальная регуляция памяти и обучения зависит от активности нейромедиаторных и нейромодуляторных церебральных механизмов, важными агентами которых являются регуляторные пептиды [8]. Однако роль пептидергической системы в организации различных функциональных состояний мозга, включая функциональное состояние в процессе обучения, недостаточно изучена.
Интегративная деятельность мозга находит свое отражение в межполушарной асимметрии. Известно, что асимметрия может проявляться на морфологическом, структурном и нейрохимическом уровнях, а ее характер зависит от гормонального статуса и функционального состояния организма [7]. Межполушарная асимметрия может претерпевать изменения при различных внешних воздействиях, что играет существенную роль в процессах памяти и обучения [7]. Высказывается предположение о неравном участии полушарий мозга и в реализации эффектов различных лекарственных веществ, включая аналоги пептидных биорегуляторов [7]. Однако нейрохимические аспекты этого вопроса практически не изучены.
Исходя из вышеизложенного, целью настоящего исследования явилось изучение влияния семакса и селанка на функциональную межполушарную асимметрию (ФМА) мозга у крыс с разным профилем моторной латерализации в условиях деструкции поля СА1 дорсального гиппокампа.
Материалы и методы исследования
Опыты выполнены на 36 самцах белых беспородных крыс массой 200-250 г, содержащихся в стандартных условиях вивария при свободном доступе к пище и воде, 12-часовом световом режиме.
Для интраназальных инстилляций применялись 1% растворы семакса и селанка. Пептиды вводились курсом (в течение 3 дней) в дозе 250 мкг/кг за 10 минут до начала эксперимента.
При определении степени межполушарной асимметрии на фоне введения семакса и селанка животные предварительно отбирались по критерию моторной латерализации. Для этого использовали следующую схему эксперимента: у животных в течение 7 дней вырабатывали условный пищедобывательный рефлекс (УПР) [5] в камере, состоящей из двух отсеков: стартового и рабочего. В рабочем отсеке располагалась площадка с лесенкой, на которой размещалась выдвижная кормушка. Животных с пищевой депривацией помещали в стартовый отсек. Через 30-60 секунд после посадки открывали дверцу стартового отсека. Звуковой сигнал служил условным раздражителем. Во время действия звукового сигнала крыса должна была подняться на площадку для пищевого подкрепления, выдвинуть кормушку за рычаг и достать из нее пищу. В этом случае решение задачи считали верным. Для анализа регистрировалось число верных решений (в процентах от числа сочетаний за один опытный сеанс). В качестве критерия выработки рефлекса выбиралось более 80% правильных реакций от числа предъявляемых сочетаний.
У крыс с выработанным УПР подсчитывали число манипуляторных движений правой и левой конечностью. Вычисляли коэффициент асимметрии (Кас), который определялся как отношение разности правосторонних (R) и левосторонних (L) манипуляторных навыков к их сумме: Кас = (R–L)/(R+L) [14]. По значению Кас животных относили к одной из групп: «правши» (0,4<Кас≤1), «левши» (–1≤Кас<–0,4) и амбидекстры (–0,4≤Кас≤0,4).
Билатеральное разрушение поля СА1 гиппокампа осуществляли электролитическим путем [2]. С этой целью крысу помещали в стереотаксический станок и в эту структуры согласно координатам атласа мозга (AP–3,3 мм, L±1,6 мм, D±2,8 мм относительно брегмы) [10], последовательно под углом 80 градусов вводили стальной электрод, изолированный на всем протяжении, кроме кончика (диаметр кончика 2 мкм). У крыс осуществляли электрокоагуляцию изучаемого отдела мозга путем пропускания постоянного тока силой 5 мА в течение 30 секунд. Хирургическое вмешательство проводили под нембуталовым наркозом (40 мг/кг).
Изучение изменения Кас у каждой из групп при трехкратном введении пептида проводили в течение 5 дней.
Оценку степени разрушения исследуемого отдела мозга изучали методом световой микроскопии с окраской фронтальных срезов мозга по Нисслю [4].
Полученные данные подвергали статистической обработке с применением программного обеспечения Statistica 6 (StatSoft, USA) с использованием t-критерия Стьюдента и дисперсионного анализа.
Результаты исследования и их обсуждение
В настоящем исследовании деструкция поля СА1 дорсального гиппокампа вызывала значительные изменения межполушарных отношений (МО), они заключались в подавлении условных моторных реакций на ранее доминирующую сторону (рис. 1).
Так, разрушение поля СА1 гиппокампа вызвало изменение Кас у крыс «правшей» на 73% в сторону уменьшения относительно исходного значения (p<0,05), у крыс «левшей» и крыс амбидекстров значение Кас увеличилось в 2,8 раза (p<0,05).
Изучение влияния селанка и семакса на изменение МО у крыс показало наличие дифференциации в их эффектах. При введении селанка особенно выражена тенденция к изменению МО у крыс «правшей». В 1 день инстилляции препарата отмечалась инверсия знака Кас, его значение было достоверно ниже в 2,3 раза относительно значения Кас после деструкции поля СА1 гиппокампа (p<0,05). Такой эффект сохранялся в течение 3 опытных дней. Уже на 4-е сутки наблюдалась тенденция в сторону дальнейшего уменьшения Кас на 42%, относительно предыдущего значения (p<0,05). На 5 день опыта показатель был равен -0,57±0,033. Это свидетельствует о приобретении животными левосторонней моторной латерализации.
У крыс «левшей» после инстилляции селанка в 1 день опыта Кас достоверно уменьшился на 39% относительно его значения в условиях деструкции поля СА1 гиппокампа (p<0,05). На 2 день опыта Кас достоверно увеличился на 39% относительно предыдущего значения (p<0,05). К 4 дню эксперимента наблюдалось достоверное уменьшение Кас на 40% (p<0,05). Так, на 5 день Кас равнялся -0,57±0,033, что свидетельствует о восстановлении изначального профиля поведения у крыс – моторные реакции преимущественно осуществлялись левой конечностью.
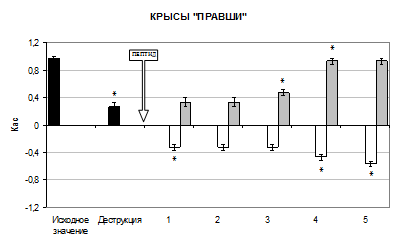
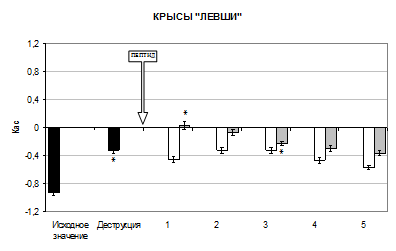
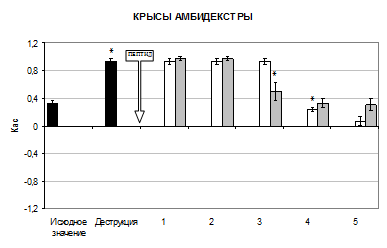
Рис. 1 Изменение Кас у крыс после разрушения поля СА1 дорсального гиппокампа при интраназальном введении пептидов (M±m, n=6). Здесь: ось абсцисс – дни опыта, ось ординат – коэффициент асимметрии, ![]() – исходное значение Кас,
– исходное значение Кас, ![]() – введение селанка,
– введение селанка, ![]() – введение семакса, * – p<0,05 относительно предыдущего значения Кас
– введение семакса, * – p<0,05 относительно предыдущего значения Кас
Селанк способствовал изменению профиля поведения у крыс амбидекстров к 4 дню опыта. Наблюдалось достоверное уменьшение Кас на 77% относительно предыдущего значения (p<0,05). На 5 день опыта Кас был равен 0,07±0,067.
Введение семакса крысам «правшам» способствовало постепенному восстановлению изначального профиля поведения, правильные условные реакции осуществлялись преимущественно на правую сторону. В 1 день опыта Кас достоверно увеличивался на 27% относительно значения показателя в условиях деструкции поля СА1 гиппокампа (p<0,05). На 3 день эксперимента Кас был выше на 42% по сравнению с предыдущим значением (p<0,05). К 4 и 5 дню эксперимента Кас был равен 0,93±0,042.
Семакс не оказывал значительного влияния на МО у крыс «левшей». Манипуляторные движения осуществлялись животными с использованием как правой, так и левой конечности.
У крыс амбидекстров влияние семакса на МО носило отсроченный характер. К 4 дню тестирования регистрировалось достоверное уменьшение Кас на 48,5% относительно предыдущего значения (p<0,05). К 5 дню Кас был ниже на 69% по сравнению со значением показателя в условиях деструкции (p<0,05).
Избирательное влияние изучаемых пептидных препаратов на деятельность правого или левого полушария головного мозга играет важную роль при смене функционального состояния организма, способствуя его адаптации к изменениям окружающей среды. Известно, что при умеренном стрессе функциональная активность чаще перемещается в субдоминантное полушарие, что сопровождается изменением центральной регуляции гомеостаза [7]. Вероятно, такое переключение является своеобразным отдыхом для деятельности доминантного полушария. Однако при некоторых видах патологии, а возможно и при нормальном старении, подобное переключение затруднено, что, по-видимому, является одним из факторов снижения качества адаптационных процессов [6, 7]. Можно предположить, что семакс и селанк оказывают свое нейропротекторное действие через восстановление способности полушарий головного мозга к такому переключению.
Таким образом, полученные в работе данные свидетельствуют о том, что проявление фармакологических эффектов семакса и селанка может осуществляться через регуляцию межполушарной асимметрии головного мозга.
Заключение
В результате исследования установлено компенсаторное действие семакса и селанка при физической деструкции поля СА1 дорсального гиппокампа.
Исследуемые пептиды отличаются по степени выраженности фармакологических эффектов. Установлены различия в их влиянии на межполушарную асимметрию у крыс с разным профилем моторной латерализации. Полученные данные подтверждают выдвинутое нами предположение о том, что одним из механизмов положительного действия пептидов на деятельность мозга является их участие в регуляции межполушарной асимметрии.
Рецензенты:
Микуляк Н.И., д.м.н., профессор, зав. каф «Физиология человека» Медицинского института ПГУ, г. Пенза;
Мельников В.Л., д.м.н., зав. кафедрой «Микробиология, эпидемиология и инфекционные болезни» Медицинского института ПГУ, г. Пенза.
Библиографическая ссылка
Латынова И.В., Купрюшин А.С., Вишнякова Ж.С. ВЛИЯНИЕ СЕМАКСА И СЕЛАНКА НА МЕЖПОЛУШАРНУЮ АСИММЕТРИЮ МОЗГА У КРЫС С РАЗНЫМ ПРОФИЛЕМ МОТОРНОЙ ЛАТЕРАЛИЗАЦИИ В УСЛОВИЯХ ДЕСТРУКЦИИ ПОЛЯ СА1 ДОРСАЛЬНОГО ГИППОКАМПА // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/ru/article/view?id=23551 (дата обращения: 04.07.2026).