



Водный гиацинт (Eichhornia crassipes (Mart.) Solms) - плейстонный гидатофит тропического происхождения из семейства понтедериевых [1,2,4,9].
Основные биологические особенности водного гиацинта:
- высокая скорость роста и вегетативного размножения при температуре воды выше +20°С, гибель при температуре воздуха ниже +4°С
- способность расти в сточных водах с концентрациями токсикантов, вызывающими гибель других водных растений.
- высокая скорость поглощения и накопления биогенов и токсикантов.
- выделение в воду биологически-активных вещества, ингибирующих развитие сине-зеленых водорослей и бактерий [1,2,3,4,5,8,9].
Эйхорния широко используется в южных регионах России для дополнительной биологической очистки коммунально-бытовых и производственных сточных вод [1-7].
Известно, что водный гиацинт светолюбив, однако влияние режима освещенности на его скорость роста, размножения и ассимиляцию биогенных элементов изучены недостаточно. Актуальность исследований связана с необходимостью оптимизации светового режима для фиторемедиации сточных вод в искусственно освещаемых закрытых бассейнах и открытых биологических прудах [1,6,7].
Эксперименты проведены на накопительных культурах в отделе биотехнологий и фиторесурсов Института морских биологических исследований РАН (г. Севастополь).
Целью работы является оценка влияния разных режимов освещенности на рост, развитие, размножение и ассимиляцию минеральных форм азота и фосфора водного гиацинта. К числу основных задач работы относится изучение влияния освещенности на рост, развитие, размножение и морфометрические признаки и оценка ассимиляционной способности эйхорнии по отношению к факторам эвтрофикации водоемов (азоту и фосфору).
Материалы и методы исследования
Растения выращивались в лаборатории (in vitro) и под открытым небом (in situ) в бассейнах объемом воды 100 л. при стартовом внесении биогенных веществ. Питательной средой являлась водная вытяжка низинного торфа (10 кг на 100 литров воды). Уровень воды в бассейнах поддерживали, компенсируя испарение воды добавлением профильтрованной водопроводной воды. Температура воды in vitro составляла в среднем 25°С и варьировала в пределах 5°С. Температура воды in situ составляла в сериях 2 и 3 в среднем 22 (16-26)°С, а в серия 4 и 5 – 28 (25-32)°С.
В качестве источника света в лабораторных условиях использовали люминисцентные лампы DRLS с освещенностью на уровне листьев растений - 3 кЛк. Проведено 5 серий эксперимента для оценки влияния светового режима (серии №1-3 - на параметры роста и размножения, № 4-5 на параметры поглощения и ассимиляции биогенов) (табл. 1).
Таблица 1
Условия проведения эксперимента с накопительными культурами водного гиацинта
|
№ серии эксперимента |
Период эксперимента, сутки |
Условия освещения |
Объем воды, л |
освещен-ность, кЛк |
рН |
|
1 |
15 |
in vitro |
50 |
3 |
8,1-9,8 |
|
2 |
15 |
in situ |
100 |
25 |
8,2-8,4 |
|
3 |
15 |
in situ |
100 |
225 |
8,3-8,8 |
|
4 |
25 |
in vitro |
50 |
3 |
8,5-9,5 |
|
5 |
25 |
in situ |
50 |
225 |
8,6-9,7 |
Продолжительность эксперимента: 15-25 суток (июнь-июль). Интенсивность освещения на уровне листовой поверхности определяли с помощью люксметра Ю-116. Ежедневно в 12 ч. измеряли физические показатели среды и раз в три дня – химические показатели среды и биологические показатели объекта исследования. Активную реакцию среды измеряли с помощью иономера И-160М.
Для определения фитомассы растения кратковременно извлекали из бассейнов и высушивали между двумя слоями фильтровальной бумаги до полного исчезновения водных следов. Измеряли морфометрические признаки листьев, подсчитывали количество дочерних раметов. Биомассу определяли с помощью аналитических весов ВЛР-200 2 класса точности с пределами измерений от 0 до 200 по ГОСТ 24104-80. Сырую фитомассу приводили к значениям абсолютно сухого вещества (АСВ) по коэффициенту 0,088. [18].
Для определения концентрации фосфора и нитратного азота отбирали порции объемом 10 мл дозатором «Biohit». Концентрацию нитратов в растворе определяли ионоселективным электродом на иономере И-160М, концентрацию фосфора – фотоэлектроколориметрическим методом Морфи-Райли.
Результаты исследования и их обсуждение
Динамика биомассы водного гиацинта в накопительной культуре соответствует модели логистического роста с линейной фазой прироста в период 3-15 сутки. Статистически значимых различий между сериями опыта с искусственным и прямым солнечным освещением за период 1-15 сутки не обнаружено (рис.1). В условиях рассеянного солнечного освещения наблюдается наименьшая скорость роста, что подтверждает светолюбивость водного гиацинта.
Наибольшие показатели чистой первичной продукции (ЧПП) и удельной ЧПП нормированной на единицу фитомассы, наблюдались в условиях прямого солнечного освещения (рис.2). Множественное попарное сравнение по критерию Краскала-Уоллиса выявило статистически значимые различия между сериями эксперимента 1 и 2 (χ²=10,0; p=0,049), а также 1 и 3 (χ²=10,6; p=0,033).
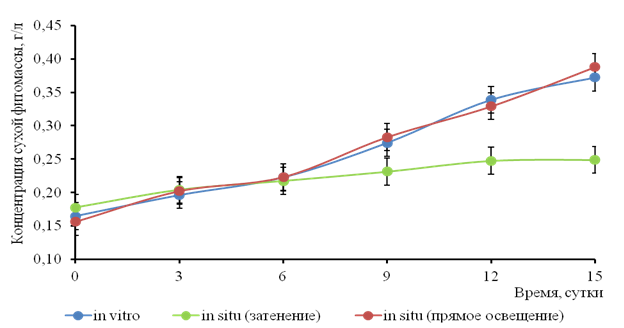
Рис. 1. Динамика АСВ водного гиацинта при культивировании при различных условиях освещения
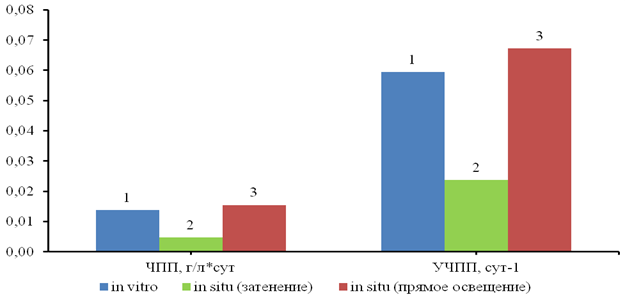
Рис. 2. Чистая первичная продукция (ЧПП, г/л*сут) и удельная ЧПП (сут-1) в накопительных культурах водного гиацинта в различных условиях освещения.
Скорость вегетативного размножения значимо различалась во всех сериях по критерию Краскала-Уоллиса (χ²=9,33; p = 0,0094), была наибольшей в условиях прямого солнечного освещения и наименьшей при затенении. Размножение при искусственном освещении in vitro шло медленнее, чем при солнечном свете in situ (рис. 3).
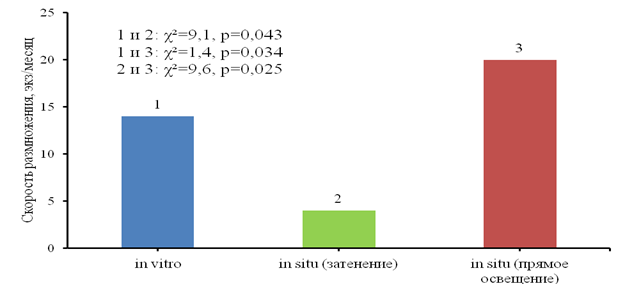
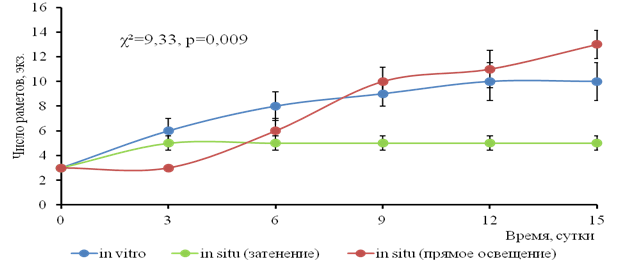
Рис. 3. Вегетативное размножение водного гиацинта при различных условиях освещения
Растения, выращенные в условиях in vitro и in situ, различались по морфометрическим характеристикам (рис.4). В лабораторных условиях они имели длинные черешки листьев; в естественных условиях на 3 сутки эксперимента черешки начали образовывать поплавки, что объясняется приспособлением к действию ветра. Удлинение черешков и образование поплавков вносит заметный вклад в суммарную листовую поверхность. Сравнения динамики длины черешков критерием Краскала-Уоллиса показало статистически значимые различия во всех сериях эксперимента (рис.5).
Суммарная площадь листовой поверхности растений максимальна при прямом солнечном освещении (0,21 м2/экз.); минимальна при затенении (0,135 м2/экз.), последнее показывает подавление вегетативного роста в условиях затенения и имеет статистически значимый характер отличия (рис.6).
а б
б
Рис. 4. Внешний вид водного гиацинта при культивировании в лабораторных (а) и естественных (б) условиях.
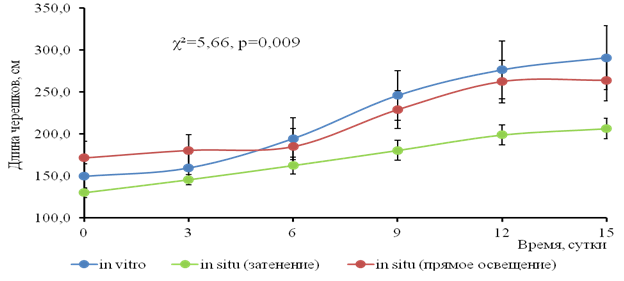
Рис. 5. Динамика длины черешка (см) водного гиацинта в культивационных бассейнах при разном режиме освещения
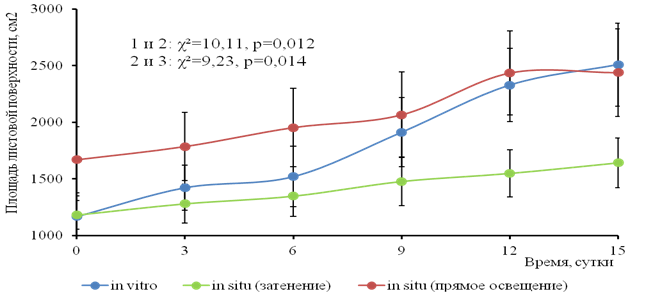
Рис. 6. Динамика площади листовой поверхности (см²) водного гиацинта при разном режиме освещения
Исследования поглощения биогенных элементов (азота и фосфора) накопительной культурой водного гиацинта в естественных и искусственных условиях прямого освещения показали следующее. Обнаружена высокая отрицательная корреляция по Спирмену биомассы с содержанием фосфора минерального (рис. 7) и умеренная корреляция (r2≥-0.6) с содержанием азота нитратного в среде. Таким образом, фосфор является ключевым лимитирующим ресурсом для вегетативного роста водного гиацинта.
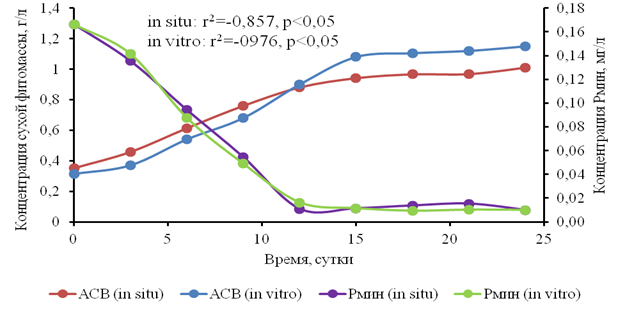
Рис. 7. Динамика концентрации минерального фосфора в культивационных бассейнах при различных условиях выращивания водного гиацинта
Функциональная зависимость чистой первичной продукции от поглощения минерального фосфора культурой водного гиацинта аппроксимируется линейной регрессией с наиболее высоким коэффициентом детерминации при солнечном освещении (табл. 2).
Таблица 2
Зависимость чистой первичной продукции водного гиацинта от поглощения минерального фосфора
|
№ эксперимента |
in situ |
in vitro |
|
Уравнение зависимости |
y = 1,659x + 0,021 |
y = 2,215x + 0,017 |
|
Коэффициент детерминации |
0,717 |
0,574 |
С помощью полученных регрессионных моделей можно прогнозировать чистую первичную продукцию на разных фазах роста культуры в зависимости от скорости поглощения минерального фосфора в условиях накопительного культивирования.
Выводы
1. В условиях рассеянного солнечного освещения наблюдается наименьшая скорость роста, размножения и чистой первичной продукции, что подтверждает светолюбивость водного гиацинта. Суммарная площадь листовой поверхности максимальна при прямом солнечном освещении и минимальна при затенении.
2. В условиях искусственного освещения отмечается удлинение листовых черешков, а в условиях in situ - развитие поплавков черешка, связанное с влиянием ветра.
3 Обнаружена высокая отрицательная корреляция фитомассы с концентрацией в воде фосфора минерального и умеренная корреляция с содержанием азота нитратного. Таким образом, фосфор является ключевым лимитирующим ресурсом для вегетативного роста водного гиацинта.
4. Функциональная зависимость чистой первичной продукции от поглощения минерального фосфора культурой водного гиацинта аппроксимируется линейной регрессией с наиболее высоким коэффициентом детерминации (R>0.7) в условиях прямого солнечного освещения.
Рецензенты:
Дмитриев А.И., д.б.н., профессор, заведующий кафедрой биологии, химии и биолого-химического образования факультета естественных, математических и компьютерных наук ФГБОУ ВО «Нижегородский государственный педагогический университет им. К. Минина», г. Нижний Новгород;
Смирнов В.Ф., д.б.н., профессор кафедры нанотехнологий и биотехнологий Института физико-химических технологий и материаловедения ФГБОУ ВО «Нижегородского государственного технического университета им. Р.Е. Алексеева», г. Нижний Новгород.
Библиографическая ссылка
Макеев И.С., Горбунова С.Ю., Коротаева М.И. ВЛИЯНИЕ ОСВЕЩЕННОСТИ НА РОСТ, РАЗМНОЖЕНИЕ И АССИМИЛЯЦИОННЫЕ ХАРАКТЕРИСТИКИ ВОДНОГО ГИАЦИНТА // Современные проблемы науки и образования. 2015. № 6. ;URL: https://science-education.ru/ru/article/view?id=23418 (дата обращения: 04.07.2026).