



Цель данной работы: изучить влияние эколого-ценотических условий на изменчивость биометрических показателей Dactylis glomerata L. в подзоне хвойно-широколиственных лесов.
Материал и методы исследований
Исследования ценопопуляций (ЦП) Dactylis glomerata проводились в разных частях Удмуртской республики (УР): центральной - (окрестности г. Ижевска), северной - (Глазовский район) и юго-западной - (Кизнерский район) в течение вегетационных сезонов 2007-2010 гг. Было изучено 15 ценопопуляция (ЦП) ежи сборной в шести типах местообитаний: I) сосновые леса; II) смешанные леса; III) пойменные луга; IV) приручьевые участки, расположенные около леса; V) суходольные луга; VI) антропогенно трансформированные экотопы (пустыри, придорожные участки, пастбище).
При описании растительных сообществ нами использованы общепринятые геоботанические методы [7, 8]. При камеральной обработке геоботанических описаний вычислялся индекс синантропизации сообществ как отношение числа синантропных видов к общему видовому составу (%). Выделение синантропных видов произведено согласно классификации Н. Г. Ильминских [4]. На основании значений коэффициента синантропизации (Кc) выделены 3 группы местообитаний в зависимости от степени антропогенной нагрузки: 1) слабой (Kc=0-15,9 %); 2) умеренной (Kc=16-30,9 %); 3) значительной (Kc≥31).
Экологическая оценка местообитаний проведена при помощи шкал Д. И. Цыганова [10] с использованием компьютерной программы «ECOSCALE» [2]. При проведении морфометрических исследований в каждой из ЦП методом случайной выборки отобрано по 3 побега с 15-20 парциальных кустов ежи сборной, находящейся в среднегенеративном возрастном состоянии (g2). У каждого из побегов проанализированы по 18 количественных параметров: высота до соцветия (Н1), длина верхнего метамера (Lmm1) и метамера, расположенного над максимально развитым листом (Lmm2), количество узлов (Nuz), длина листового влагалища (Lv), длина (L) и ширина (W) листовой пластинки, высота соцветия (Lsoc), количество веточек первого порядка (Nvet), у последних - длина двух нижних (L1 и L2) и расстояние между ними (L3). На основании полученных данных вычислены: общая высота побега (H=Н1+Lsoc), площадь листовой пластинки (S=2/3LW) и другие аллометрические показатели [3] - H/Lmm1, Н1/Lsoc, L/W, L/Lv.
Статистический анализ результатов исследований проведен в соответствии с общепринятыми стандартными методами описательной статистики с использованием компьютерной программы «STATISTICA 5,5» и «MS EXCEL».
Результаты исследования и их обсуждение
Для изучения влияния эколого-ценотических условий на биометрические показатели Dactylis glomerata мы использовали результаты исследований геоботанических описаний растительных сообществ и балльных оценок шкал Д. Н. Цыганова [10] (табл. 1).
Таблица 1
Характеристика фитоценозов с участием Dactylis glomerata L. (Удмуртская республика, 2007-2010 гг.)
|
Типы место- обитаний |
№ п/п
|
Ассоциации
|
Балльные оценки местообитаний по экологическим шкалам* Д.И. Цыганова [10] |
|||||
|
Hd |
Tr |
Nt |
Rc |
fH |
Lc |
|||
|
I |
1 |
Pinus sylvestris+Carex rhizina+ Fragaria vesca+Dactylis glomerata |
12,02 |
6,57 |
6,16 |
6,81 |
5,58 |
3,6 |
|
2 |
Pinus sylvestris+ Poa angustifolia Dactylis glomerata |
11,51 |
6,58 |
5,63 |
6,73 |
5,55 |
3,45 |
|
|
3 |
Pinus sylvestris+Oxalis acetosella+ Fragaria vesca+Dactylis glomerata |
11,65 |
6,74 |
5,92 |
6,93 |
5,89 |
3,47 |
|
|
II |
4 |
Picea abiesXfennica+Betula pendula +Calamagrostis epigeios+ Dactylis glomerata |
12,53 |
6,27 |
6,12 |
7,01 |
5,7 |
3,92 |
|
5 |
Picea abiesXfennica+Betula pendula+ Aegopodium podagraria+ Dactylis glomerata |
12,4 |
6,72 |
6,29 |
6,76 |
5,7 |
3,68 |
|
|
III |
6 |
Achillea millefolium+Poa pratensis+ Calamagrostis epigeios |
12,1 |
7,93 |
6,2 |
7,43 |
6,14 |
2,86 |
|
7 |
Dactylis glomerata+Bromopsis inermis |
12,94 |
8,36 |
6,79 |
6,91 |
5,79 |
2,89 |
|
|
IV
|
8 |
Tussilago farfara+Dactylis glomerata |
11,71 |
7,41 |
6,27 |
7,38 |
5,81 |
3,19 |
|
9 |
Filipendula ulmaria+ Dactylis glomerata |
12,4 |
7,36 |
6,56 |
7,44 |
5,96 |
3,21 |
|
|
10 |
Filipendula ulmaria+ Dactylis glomerata+ Urtica dioica |
11,3 |
7,63 |
6,89 |
7,14 |
5,87 |
3,16 |
|
|
V |
11 |
Achillea millefolilium+Taraxacumofficinale+ Dactylis glomerata |
11,59 |
7,13 |
5,63 |
6,88 |
5,72 |
2,99 |
|
12 |
Festuca pratensis+ Poa pratensis+ Dactylis glomerata |
11,47 |
7,61 |
5,93 |
7,3 |
6,03 |
2,81 |
|
|
VI |
13 |
Calamagrostis epigeios+ Poa angustifolia+ Dactylis glomerata |
10,46 |
7,93 |
5,61 |
7,11 |
6,43 |
2,54 |
|
14 |
Dactylis glomerata+Poa pratensis+ Trifolium pratense |
11,25 |
7,74 |
5,57 |
6,78 |
5,49 |
2,71 |
|
|
15 |
Cichorium intybus+ Tanacetum vulgare+ Dactylis glomerata |
10,89 |
7,52 |
5,85 |
6,87 |
6,12 |
2,76 |
|
Примечание: экологические шкалы Д. И. Цыганова: Hd - увлажнения почв, Tr - солевого режима почв, Nt - богатства почв азотом, Rc - кислотности почв, Lc - освещенности-затенения, fH - переменности увлажнения; № 1-15 - порядковый номер ЦП.
Изученные сообщества (ЦП 1-15) Dactylis glomerata характеризуются значительным разнообразием луговых, сорно-рудеральных и лесо-луговых растений. В исследуемых ценозах данный вид, чаще всего, является доминантом, реже ассектатором с проективным покрытием (ПП) 15-75 %. Чаще всего сопутствующими видами Dactylis glomerata являются: Poa pratensis L., Achillea millefolium L., Festuca pratensis Huds., Veronica chamaedrys L., Taraxacum officinale Wigg. s. l и т.д.
Лесные сообщества (ЦП № 1-5) характеризуются слабой (Кc=5,5-13,1 %) и умеренной (Кc=21,0-22,9 %) антропогенной нарушенностью. В травянистом ярусе сосновых лесов (ЦП 1-3) доминируют Oxalis acetosella L., Carex rhizina Blyttex Lindblom, Fragaria vesca L.; в смешанных лесах (ЦП 4-5) - Aegopodium podagraria L. Проективное покрытие (ПП) ежи сборной составляет 25-55 %. Ценопопуляции Dactylis glomerata L., приуроченные к лесным опушкам, представлены редко расположенными парциальными кустами.
Луговые сообщества (ЦП 6-7, 11-12) и приручьевые участки (ЦП 8-10) также характеризуются слабой (Кc=12,0 %) и умеренной антропогенной нарушенностью (Кc=17-30,3 %), травостой некоторых из них (ЦП 7, 12) подвержен ежегодному сенокошению. Наряду с Dactylis glomerata L., в пойменных лугах (ЦП 6-7) и на приручьевых участках (ЦП 8-10) обильно представлены Ranunculus acris L., Bromopsis inermis (Leyss.) Holub, Filipendula ulmaria (L.) Maxim., Geranium pratense L.; на суходольных лугах (ЦП 11-12) - Achillea millefolium L., Taraxacum officinale Wigg. s. l., Poa pratensis L., Festuca pratensis Huds. Проективное покрытие ежи сборной на луговых сообществах составляет 50-75 %, в приручьевых участках - 30-35 %.
Ценопопуляции (ЦП 8-10) Dactylis glomerata приручьевых участков на территории Удмуртии встречаются довольно редко. Они расположены у подножия холмов, вдоль ручьев, в кустарниковых зарослях, близ лесных участков, где откладываются аллювиальные и делювиальные отложения. Растения Dactylis glomerata L. из указанных ценопопуляций отличаются наибольшей высотой побегов.
Антропогенно трансформированные местообитания характеризуются уплотненным субстратом, содержащим строительный и бытовой мусор. В таких условиях экотопа сообщества (ЦП 13-15) характеризуются значительным уровнем синантропизации (Кс = 31-37 %), низкими значениями проективного покрытия Dactylis glomerata L. (15-25 %) и бедным видовым разнообразием, состоящим в основном из луговых и сорно-рудеральных растений: Artemisia vulgaris L., Arctium tomentosum L., Trifolium pratense L., Tanacetum vulgare L., Cichorium intybus L., Achillea millefolium L., Pimpinella-saxifraga L., Plantago major L. [1].
На основании анализа шкал Д. Н. Цыганова (табл. 1), характеризующих эколого-ценотические условия растительных сообществ, нами выявлена взаимосвязь между балльными оценками исследуемых режимов (Hd, Tr, Nt, Rc, Lc, fH) и биометрическими показателями побегов Dactiylis glomerata L. при помощи корреляционного анализа (табл. 2). В результате установлена значимая прямо пропорциональная зависимость большинства признаков, характеризующих размеры листа (L,W, Lv, S, L/W L/Lv) и стебля (Н, Lmm2, H1, H/Lmm1) от факторов увлажнения почв (Hd) и освещенности - затенения (Lс) местообитаний, и обратно пропорциональная (Nuz, L, L/W L/Lv) - от солевого режима почв (Tr).
Таблица 2
Корреляционная зависимость между морфологическими параметрами побегов Dactylis glomerata и балльными оценками экологических шкал Д. Н. Цыганова (Удмуртская республика, 2007-2010 гг.)
|
Морфологические признаки |
Экологические шкалы Д. Н. Цыганова (1985) |
|||||
|
Hd |
Tr |
Nt |
Rc |
fH |
Lс |
|
|
H |
0,53** |
-0,22 |
0,77*** |
0,34 |
-0,12 |
0,54** |
|
H1 |
0,43 |
-0,31 |
0,68*** |
0,31 |
-0,14 |
0,54** |
|
Nuz |
0,62*** |
-0,52* |
0,23 |
-0,12 |
-0,22 |
0,65** |
|
L |
0,43 |
-0,51* |
0,48 |
-0,05 |
-0,3 |
0,78*** |
|
W |
0,58* |
0,26 |
0,77*** |
0,51* |
0,07 |
0,19 |
|
Lmm1 |
0,18 |
0,06 |
0,27 |
0,23 |
0,26 |
-0,07 |
|
Lv |
0,57* |
-0,35 |
0,75** |
0,16 |
-0,21 |
0,7** |
|
Lmm2 |
0,52* |
-0,06 |
0,81*** |
0,4 |
0,08 |
0,38 |
|
Lsoc |
0,65** |
0,004 |
0,91*** |
0,39 |
0,01 |
0,46 |
|
L1 |
0,6** |
0,14 |
0,87*** |
0,44 |
0,06 |
0,32 |
|
L2 |
0,6** |
0,12 |
0,88*** |
0,42 |
0,08 |
0,33 |
|
L3 |
0,59* |
-0,03 |
0,9*** |
0,35 |
0,03 |
0,49 |
|
Nvet |
0,42 |
-0,01 |
-0,002 |
-0,35 |
-0,01 |
0,16 |
|
H/Lmm1 |
0,59* |
-0,47 |
0,45 |
0,06 |
-0,21 |
0,76*** |
|
H1/Lsoc |
-0,04 |
-0,48 |
-0,12 |
0,16 |
0,02 |
0,23 |
|
L/W |
0,50* |
-0,711*** |
0,38 |
-0,13 |
-0,341 |
0,9*** |
|
L/Lv |
0,36 |
-0,76*** |
0,21 |
-0,03 |
-0,3 |
0,8*** |
|
S |
0,50* |
-0,29 |
0,72*** |
0,28 |
-0,11 |
0,64** |
Примечание: уровни значимости: * - p<0,05, ** - p<0,01, *** - p<0,001. Условные обозначения морфологических признаков и балльных оценок Д. Н. Цыганова приведены выше в тексте.
К условиям богатства почв азотом (Nt) наиболее чувствительны параметры соцветия (Lsoc, L1, L2, L3), листа (L, W, S) и высоты побега. Указанные признаки, согласно результатам однофакторного дисперсионного анализа, вносят наибольший вклад в межпопуляционную изменчивость ежи сборной, что позволяет использовать их в качестве интегральных показателей при оценке эколого-ценотических условий местообитаний ежи сборной.
Влияние эколого-ценотических условий на биометрические показатели Dactylis glomerata также подтверждены результатами однофакторного дисперсионного анализа, который позволил установить достоверные различия (p<0,05) между морфометрическими параметрами побегов ежи сборной на разных типах местообитаний. Их усредненные показатели представлены в табл. 3.
Таблица 3
Морфологическая характеристика ценопопуляций Dactylis glomeratа на разных типах местообитаний (Удмуртская республика, 2007-2010 гг.)
|
Морфологические признаки |
I |
II |
III |
IV |
V |
VI |
|
M±m |
M±m |
M±m |
M±m |
M±m |
M±m |
|
|
H |
110,6±0,74 |
129,5±1,07 |
115,5±1,03 |
155,2±1,48 |
114,2±0,87 |
80,6±0,97 |
|
H1 |
94,5±0,74 |
116,4±0,99 |
101,9±0,95 |
140,5±1,37 |
103,8±0,85 |
72,3±0,9 |
|
Nuz |
6,1±0,03 |
5,8±0,04 |
5,8±0,04 |
5,3±0,08 |
5,5±0,05 |
5,2±0,03 |
|
Lmm1 |
18,5±0,27 |
21,9±0,49 |
22,7±0,25 |
24,4±0,75 |
25,6±0,38 |
19,2±0,29 |
|
Lmm2 |
17,3±0,20 |
23,2±0,32 |
22,2±0,32 |
26,5±0,55 |
20,8±0,44 |
16,9±0,32 |
|
Lv |
15,5±0,13 |
16,4±0,17 |
15,6±0,22 |
18,6±0,43 |
12,4±0,15 |
10,2±0,12 |
|
L |
34,5±0,3 |
33,6±0,40 |
26,6±0,27 |
33,5±0,42 |
22,1±0,26 |
17,3±0,26 |
|
W |
0,76±0,01 |
0,89±0,02 |
0,94±0,01 |
0,93±0,01 |
0,81±0,01 |
0,75±0,01 |
|
S |
17,8±0,26 |
19,6±0,36 |
16,7±0,24 |
20,9±0,37 |
12,1±0,22 |
8,82±0,18 |
|
Lsoc |
11,7±0,14 |
13,04±0,23 |
13,6±0,20 |
14,7±0,27 |
10,4±0,17 |
8,35±0,14 |
|
L1 |
5,5±0,08 |
6,45±0,12 |
7,07±0,13 |
7,3±0,14 |
5,3±0,11 |
4,2±0,08 |
|
L2 |
3,9±0,06 |
4,58±0,09 |
5,03±0,11 |
5,2±0,12 |
3,8±0,09 |
3,02±0,07 |
|
L3 |
3,5±0,05 |
3,77±0,06 |
3,9±0,06 |
4,2±0,09 |
2,9±0,05 |
2,7±0,05 |
|
Nvet |
14,5±0,08 |
14,64±0,10 |
15,3±0,09 |
12,5±0,12 |
13,9±0,13 |
14,1±0,09 |
|
H/Lmm1 |
7,1±0,13 |
6,59±0,15 |
5,2±0,07 |
6,2±0,19 |
4,6±0,08 |
4,3±0,07 |
|
H1/Lsoc |
8,9±0,11 |
9,58±0,18 |
7,8±0,1 |
10,4±0,21 |
10,5±0,18 |
9,1±0,15 |
|
L/W |
46,0±0,53 |
39,11±0,49 |
28,9±0,34 |
36,7±0,57 |
27,4±0,4 |
23,2±0,35 |
|
L/Lv |
2,3±0,02 |
1,97±0,02 |
1,8±0,02 |
2,0±0,03 |
1,8±0,03 |
1,7±0,02 |
Примечание: тип местообитаний: I - сосновые леса; II - смешанные леса; III - пойменные луга; IV- приручьевые участки, расположенные около леса; V- суходольные луга; VI - антропогенно трансформированные экотопы; M - среднее значение морфологического параметра, m - ошибка среднего значения. Условные обозначения морфологических признаков приведены выше в тексте.
ЦП ежи сборной, расположенные в приручьевых участках (ЦП 8-10) и антропогенно трансформированных местообитаниях (ЦП 13-15), значимо отличаются от всех исследованных ценопопуляций по большинству параметров (H, H1, Lmm2, Lv, S, L/W, L/Lv). В первом случае (ЦП 8-10), наблюдаются максимальные средние значения большинства морфологических признаков, во втором (ЦП 13-15) - наименьшие, что обусловлено значительной степенью антропогенной нагрузки.
В лесных и приручьевых сообществах отмечены максимальные значения параметров листа (L, L/Lv, L/W), что вероятно, является адаптационным механизмом увеличения фотосинтезирующей поверхности в условиях полутени (Lс=3,19-3,6). ЦП лесных сообществ значимо (p<0,05) отличаются от других ЦП по высоте побега (H)и некоторым параметрам листовой пластинки (Lv, L/W, L/Lv), за исключением некоторых ЦП, расположенных в пойменных лугах (ЦП 6), суходольных лугах (ЦП 11-12) и приручьевых участках (ЦП 9-10).
Средние размеры растений ежи сборной (H=114,2-115,2 см) наблюдаются на луговых участках, что, может быть связано, с повышенной плотностью побегов фитоценозов в данных местообитаниях. ЦП, расположенные в указанных экотопах, значимо отличаются от всех остальных ЦП по высоте побега и некоторым параметрам листа (L, L/W) [5].
Результирующий эффект адаптивного морфогенеза проявляется через становление корреляционной структуры и тесноты связей между признаками организма в популяциях, изменчивость которых в зависимости от условий среды может служить индикатором внешних воздействий [9].
Для определения тесноты связей между морфологическими параметрами использовали значения коэффициента корреляции Спирмена (Rs) (рис. 1), который позволил установить преобладающее количество слабых связей (0,3
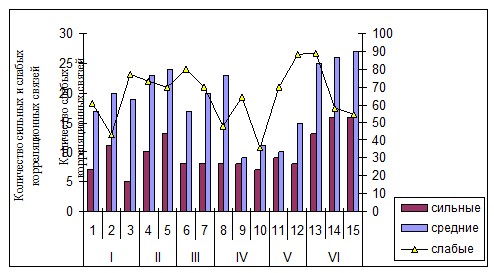
Рис. 1. Частота встречаемости достоверных сильных, средних и слабых корреляционных связей между морфологическими параметрами Dactylis glomerata (Удмуртская республика, 2007-2010 гг.)
Примечание: I) сосновые леса; II) смешанные леса; III) пойменные луга; IV) приручьевые участки, расположенные около леса; V) суходольные луга; VI) антропогенно трансформированные сообщества; 1-15 - номера ценопопуляций.
Частота встречаемости сильных корреляционных связей (0,7
Таким образом, число и теснота корреляционных связей морфологических признаков ежи сборной может меняться в зависимости от экологических условий местообитаний, что важно при изучении адаптационных механизмов указанного вида к условиям среды.
Выводы:
1. Выявленная амплитуда шкал по Д. Н. Цыганову по ряду экологических режимов (Lc, Nt, Tr, Hd) свидетельствует о высоких адаптационных возможностях Dactylis glomerata. Наибольшее влияние на изменчивость биометрических показателей оказывают условия увлажнения, освещенности-затенения и богатства почв азотом, что подтверждено результатами корреляционного анализа (p<0.05).
2. Морфологические параметры Dactylis glomerata статистически достоверно отличаются в разных типах экотопов (p<0,05). Максимальными средними значениями большинства морфологических параметров характеризуются растения Dactylis glomerata, произрастающие на приручьевых участках, минимальными - на антропогенно трансформированных местообитаниях.
3. Корреляционные связи морфологических параметров ежи сборной изменяются в зависимости от экологических условий местообитаний. Реакция на стресс данного вида проявляется в увеличении числа и тесноты связей в условиях повышенной антропогенной нагрузки, что может быть показателем при проведении биоиндикационных исследований.
4. При проведении биоиндикационных и мониторинговых исследований местообитаний Dactylis glomerata в качестве информативных показателей рекомендуем использовать параметры, характеризующие лист (L, Lv, S, L/W L/Lv) и стебель (H, H/Lmm1).
Работа выполнена при поддержке НОЦ «Мониторинг окружающей среды и прогнозирование состояния биосферы» по научному направлению «Рациональное природопользование» в рамках ФЦП «Научные и научно-педагогические кадры инновационной России» на 2009-2013 годы.
Рецензенты:
Ильминских Н.Г., д.б.н., профессор кафедры общей биологии ФГБОУ ВПО «Государственный аграрный университет Северного Зауралья», г. Тюмень, заведующий лабораторией УРАН «Тобольская комплексная биологическая станция УРО РАН», г. Тобольск.
Туганаев В.В., д.б.н., профессор кафедры общей экологии ФГБОУ ВПО «Ижевский государственный университет», г. Ижевск.
Библиографическая ссылка
Красноперова С.А. ВЛИЯНИЕ ЭКОЛОГО-ЦЕНОТИЧЕСКИХ УСЛОВИЙ НА ИЗМЕНЧИВОСТЬ БИОМЕТРИЧЕСКИХ ПОКАЗАТЕЛЕЙ DACTYLIS GLOMERATA L. В ПОДЗОНЕ ХВОЙНО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ // Современные проблемы науки и образования. 2015. № 5. ;URL: https://science-education.ru/ru/article/view?id=22152 (дата обращения: 26.06.2026).