



Цель работы – оценить морфофункциональное состояние шишковидной железы рыжей полёвки с учётомпродолжительности светлого времени суток и численности при клеточном содержании в виварии.
Материал и методы исследования
Работавыполнена на 13-тидвухмесячныхсеголеткахвесенне-летней и позднелетней генерацийв соответствии с «Правилами проведения работ с использованием экспериментальных животных» (1977). Детёныши-самцы, рождённые от изъятых из природной среды самок,содержались при естественном освещении и выводились из экспериментав 1000-1130ч путём декапитации. После отсепаровывания кожи, перерезки в области лобного шва, симметричного кругового вскрытия в направлении от лобной части к затылочной, отламывания костей свода черепа шишковидная железа отделялась от мозговых оболочек. На 2 ч выделенный материалпогружался в смесь 4% параформальдегида и 2,5% глютаральдегида на 0,1М какодилатном буфере (рН 7,4, 4⁰С),в течение 2 ч постфиксировался в 1%OsO4 на 0,1М какодилатном буфере (рН 7,4, 4⁰С), обезвоживался в этиловом спирте и заключалсяв смесь смол эпон–аралдит. Срезы изготавливались наультратоме «LeicaEMUC7» (Австрия). Полутонкие срезы, окрашенные азуромII, изучали в микроскопе «PrimoStar» («CarlZeiss», Германия) с цифровой фотокамерой G-10 («Саnon», Япония) и программным обеспечением «AxioVision» («CarlZeiss», Германия).В каждом срезе в 10-ти полях площадью 0,016 мм2 измерялиядра, ядрышкипинеалоцитов.Ультратонкие срезы, контрастированныеуранилацетатом и цитратом свинца, исследовали втрансмиссионном микроскопе «JEM-100 СХ II» (JEOL, Япония). С помощью тест-системы,соответствующей 47 мкм2среза при увеличении ×25000,в 10-ти случайно отобранных пинеалоцитахизмерялиудельный объём органелл. Данные морфометрии обрабатывали с помощью пакета программ«StatisticaforWindows», версия 6.1 (StatSoftInc., США). Для кариометрических показателей вычислялисреднее значениеМ,стандартное отклонениеs,оценивали значимость различий поt-критерию Стьюдента. Для органеллометрических показателей вычисляли медиануМе,25%-й и 75%-й квартили. Значимостьразличий оценивали поU-критерию Манна–Уитнина уровне p<0,05.
Результаты исследования и их обсуждение
У рыжей полёвки шишковидная железа имеет целлюлярный тип строения. ПинеалоцитытипаI (ранее называлисветлыми) располагаются небольшими группами и одиночно среди пинеалоцитовтипаII (ранее называлитёмными). Перикарионы и отросткипинеалоцитовтесно прилегают друг к другу,слабо, с разной степенью выраженности окрашиваются азуром II. Между пинеалоцитамии в перикапиллярном пространстве обнаруживаются псевдовакуоли с конкрециями.Ядрапинеалоцитоввытянуты в различных направлениях, имеют 1–2 ядрышка, мелкодисперсныйхроматин, складчатуюоболочку. Митохондрии придаютцитоплазме зернистый характер. Ядра пинеалоцитов типа II более гиперхромные, содержат большемелких глыбокгетерохроматина, чем клетки типа I. В пинеалоцитах типа Iа располагаются комплексы параллельных цистерн гранулярной эндоплазматической сети. Впинеалоцитах типа Ib – стопки Гольджи и одиночные цистерны гранулярной эндоплазматической сети. Во всех светлыхклетках выявляются секреторные везикулы,синаптические сферы, ленты. Дляпинеалоцитов типаIb характернанейрокриннаясекреция, при которой секреторные везикулы диаметром 130–160 нм с плотной сердцевиной и узким светлым ореолом, производные комплексаГольджи, возможно, транспортирующиеантигонадотропный пептид и белок-носитель мелатонина,освобождаются от содержимогопо мерокринному способу в булавовидных окончаниях отростков. Гормоны поступают в кровь, белок-носитель кальцинируется, формируя отложения мозгового песка в перикапиллярном пространстве и псевдовакуолях между пинеалоцитами. В клетках типаIа находит место эпендимоподобная секреция, при которойсекреторных везикул, производные гранулярной эндоплазматической сети,диаметром 180–450 нм, содержащие хлопьевидный материал,отпочковываются от перикариона.
Усеголеток рыжей полёвки индивидуального содержания при укорочении суточной фотофазы вшишковидной железевозрастает число и размеры псевдовакуолей с конкрециями. У совместно содержавшихся сеголеток весенне-летней генерациив пинеалоцитах типаIb увеличиваются стопки Гольджи и митохондрии. Митохондриальныекристы широкие, митохондриальный матрикс частично просветлён, что свидетельствует об активизации секреторной деятельностипинеалоцитов. Гранулярные везикулы выявляются не только в булавовидных терминалях отростков пинеалоцитов, но и на периферии перикарионов вблизи псевдовакуолей с мембранами и конкрециями, то есть их содержимое отделяется не только мерокринным способом, нои почкованием (рис. 1).
При совместном содержании и укорочении фотофазы соотношение пинеалоцитов светлые : тёмные сдвигается в сторону последних. Пинеалоциты типа II имеют крупные ядрышки, многочисленные свободные рибосомы и полисомы в цитоплазме. Цистерны гранулярной эндоплазматической сети с узким просветом. Обнаруживаются единичные везикулы Гольджи. По-видимому, тёмные пинеалоциты становятся «резервными» несекретирующими гормоныклетками. Об активизации функционирования шишковидной железы у рыжей полёвки при укорочении суточной фотофазы и моделировании стресса на высокую плотность популяции свидетельствует явление гипертрофиисветлыхпинеалоцитов. Их ядра, благодаря гигантским продольным складкам оболочки – «кариосомам», уподобляются кофейным зёрнам. Псевдовакуоли с конкрециями обнаруживаются вблизи перикарионов. Астроциты нередко располагаются парами или формируют более выраженные скопления, что свидетельствует о пролиферации и разрастании астроглии(рис. 2).
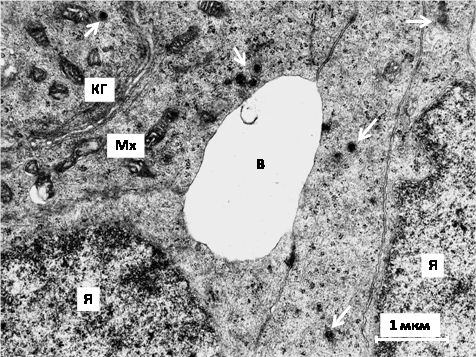
Рис. 1. Фрагмент шишковидной железы рыжей полёвки весенне-летней генерации совместного содержания: КГ – комплекс Гольджи, Мх – митохондрии, Я – ядра пинеалоцитов, В – межклеточная «вакуоль», стрелки – секреторные везикулы в перикарионе и отростке пинеалоцитов. Электронограмма
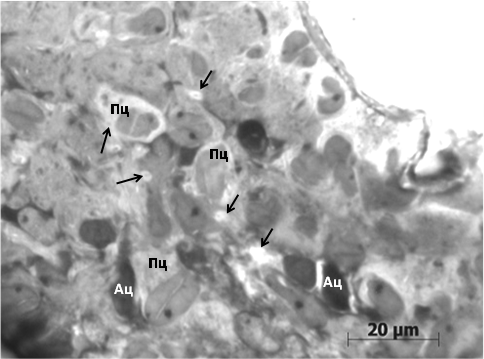
Рис. 2. Фрагент шишковидной железырыжей полёвкипозднелетней генерации совместного содержания: Ац – астроциты, Пц – пинеалоциты, стрелки – межклеточные псевдовакуоли. Окраска азуром II.
Оценка комбинированного эффектадействия высокой плотности популяции и естественного укорочения светлого времени суток на функциональное состояние шишковидной железыдвухмесячных самцов рыжей полёвки по кариометрическим и органеллометрическим показателям светлых пинеалоцитов типа Ib с использованием уравнений Уэбба свидетельствует о синергическом влиянии двух факторов на нейросекрецию.У самцов позднелетней генерации, содержавшихся совместно, синергизм комбинированного действия проявляется в отношении возрастания удельного объёмакомплекса Гольджи и митохондрий в цитоплазме пинеалоцитов типа Ib. Антистрессорный фотопротективный эффект (антагонизм комбинированного действия укорочения фотофазы и высокой плотности популяции)проявляется в отношении изменения морфометрических параметров компонентов белоксинтезирующего аппаратапинеалоцитов типа Ib – диаметра ядрышек и удельного объёма гранулярной эндоплазматической сети в цитоплазме, что, возможно, связано с обеспечением функционирования органа на следующий год после рождения(таблица).
Кариометрические показатели (M±s) и органеллометрические (Ме (Q1–Q3), р<0,05) Ib-пинеалоцитоврыжей полёвки при укорочении фотофазы и высокой плотности популяции
|
Содержание |
Совместное (С) |
Индивидуальное (И) |
Эффект комби- нирован- ного действия |
||
|
Генерация, численность |
ПЛ – позднелетняя(n=4) |
ВЛ –весенне-летняя(n=3) |
ПЛ – позднелетняя(n=3) |
ВЛ –весенне-летняя(n=3) |
|
|
индекс |
i1,2=1-ПЛС/ВЛИ |
i2=1-ВЛС/ВЛИ |
i1=1-ПЛИ/ВЛИ |
||
|
SЯ, мкм2 |
53,4±12,8
|
58,6±12,8 |
43,4±19,6 |
55,8±2,28 |
антагонизм |
|
iSя |
0,032 |
0,050 |
0,004 |
||
|
dЯк, мкм |
1,47±0,13
|
1,44±0,18 |
1,41±0,24 |
1,44±0,06 |
антагонизм |
|
idяк |
0,007 |
0 |
0,104 |
||
|
VГЭС, % |
1,97 (1,87–2,03) |
3,70 (2,81–4,86) |
2,22 (1,58–2,42) |
2,60 (2,43–2,95) |
антагонизм |
|
iVгэс |
0,268 |
0,375 |
0,175 |
||
|
VМх, % |
5,18 (4,35–6,42) |
7,92 (7,48–8,29) |
6,06 (5,24–6,37) |
6,87 (6,81–7,42) |
синергизм |
|
iVмх |
0,273 |
0,111 |
0,150 |
||
|
VКГ, % |
2,37 (2,24–3,23) |
4,89 (4,65–5,16) |
2,94 (1,66–3,24) |
4,58 (4,31–4,74) |
синергизм |
|
iVкг |
0,477 |
0,079 |
0,351 |
||
Примечание. SЯ – площадь ядра, dЯк – диаметр ядрышка, VГЭС – удельный объём гранулярной эндоплазматической сети, VМх – митохондрий, VКГ – комплекса Гольджи. ПЛИ, ВЛСи ПЛС – доля ВЛИ(%). Антагонизм: i1,2<(i1+i2)-(i1×i2). Синергизм: i1,2>(i1+i2)-(i1×i2).
Заключение
Таким образом, шишковидная железа удвухмесячных сеголеток-самцов рыжей полёвки имеет вариабельную программу функционирования, проявляяморфологические признаки усилениясекреторной активностии широкое многообразие способов секреции при моделировании высокой плотности популяции. Укорочение светлого времени суток в комбинации со стрессом на высокую плотность популяции усиливает сдвиг величины морфометрических показателей активизации секреции пинеальных гормонов, сдерживающих половое созревание, и оказывает благоприятное влияние на структурные преобразования паренхимы, связанные с обеспечением функционирования шишковидной железы на следующий год после рождения.
Рецензенты:
Алябьев Ф.В., д.м.н., профессор, заведующий кафедрой судебной медицины с курсом токсикологической химии ГБОУ ВПО «Сибирский государственный медицинский университет» Министерства здравоохранения РФ, г. Томск;
Мустафина Л.Р., д.м.н., профессор кафедры гистологии, эмбриологии и цитологии ГБОУВПО «Сибирский государственный медицинский университет» Министерства здравоохранения РФ, г. Томск.
Библиографическая ссылка
Герасимов А.В., Костюченко В.П., Логвинов С.В., Денисов А.А., Потапов А.В., Варакута Е.Ю. МОРФОФУНКЦИОНАЛЬНОЕ СОСТОЯНИЕ ШИШКОВИДНОЙ ЖЕЛЕЗЫ РЫЖЕЙ ПОЛЁВКИ ПРИ УКОРОЧЕНИИ СУТОЧНОЙФОТОФАЗЫ И ВЫСОКОЙ ПЛОТНОСТИ ПОПУЛЯЦИИ // Современные проблемы науки и образования. 2015. № 3. ;URL: https://science-education.ru/ru/article/view?id=20113 (дата обращения: 01.07.2026).